Pharyx ( two bulbi without intermediate piece ): 15 µm - 16 µm
Diameter of the mouth ( narrow, extended, terminal ): unknown
Dorsal scales: 12 - 16 thick, curved, simple and immobile spines; of which 3 dorsally (29-30 µm), 9-13 laterally, extending around the posterior end(18-21 µm)
Ventral scales: two ciliated bands, united anteriorly, visible dorsally and laterally
Diameter of the mouth ( elliptical, two teeth (9x5 µm) ): 8 x 20 µm
Dorsal scales: no scales, but closed cuticular layer
Ventral scales: broad hypostomium with transverse furrow (38-42 µm); two rows of cilia; additionally two ciliary tufts on head (median), one pair of ciliary tufts laterally on head
Oecology: Psammon (sandy ground)
Similar species: Similar to Ichthydium, but completely covered with structureless cuticle (in Ichthydium the cuticle is usually finely structured); well demarcated by complex structure of mouth.
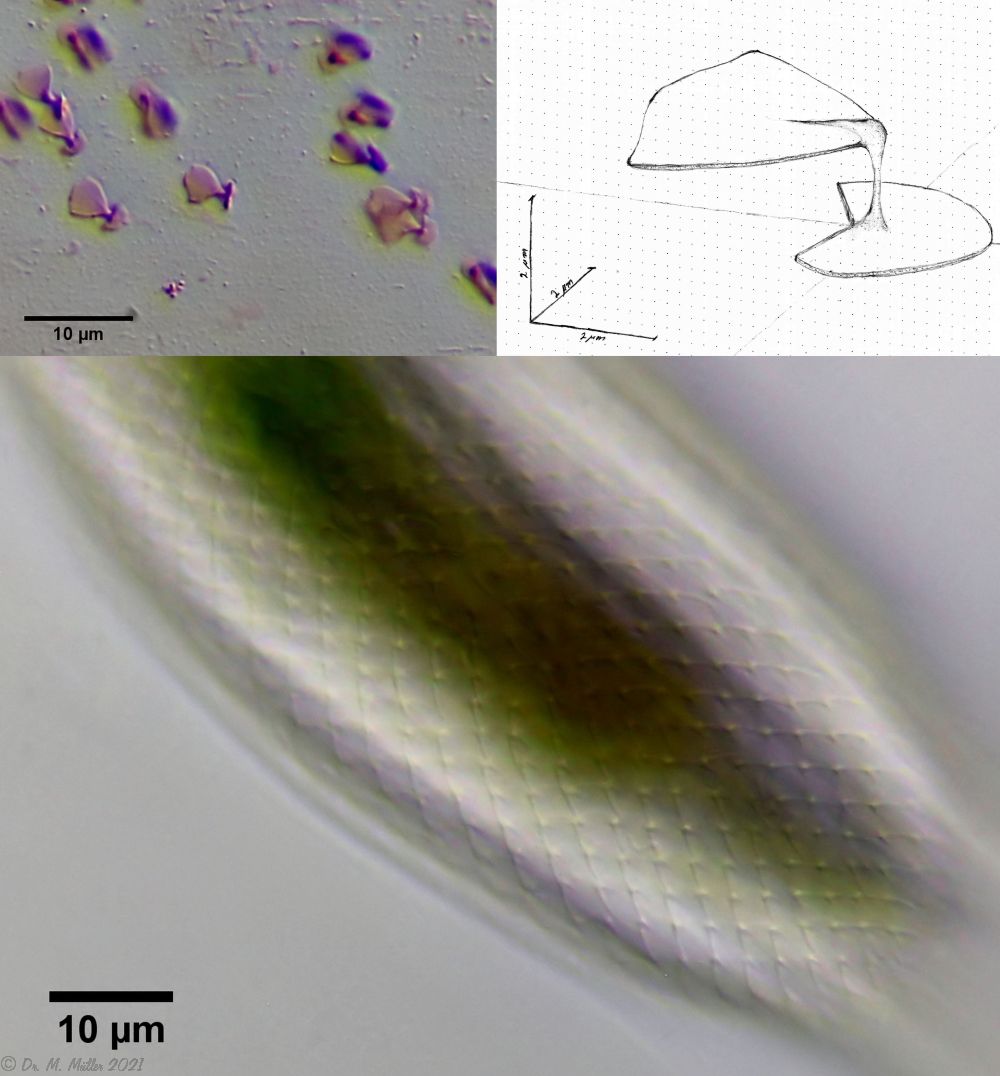
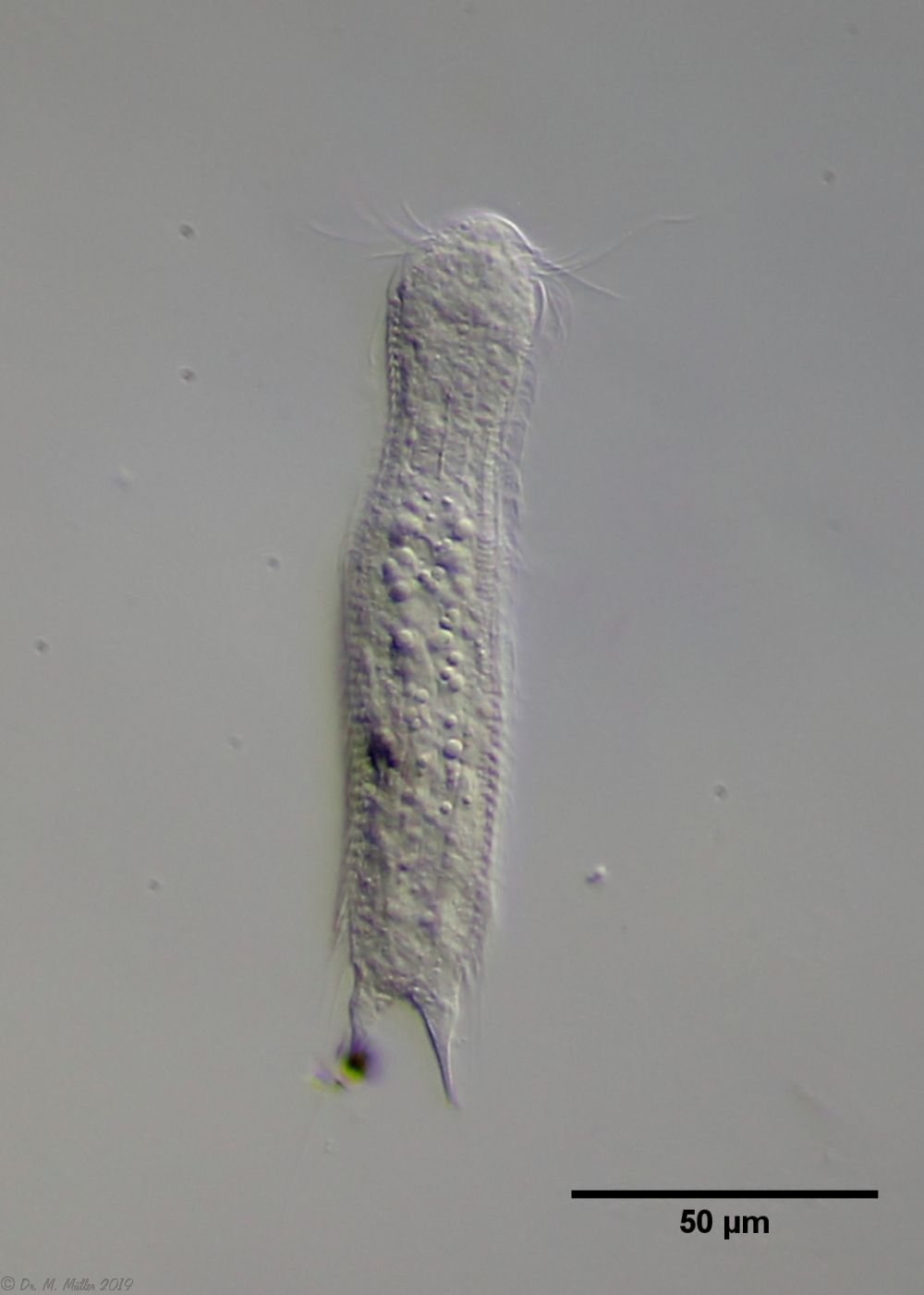
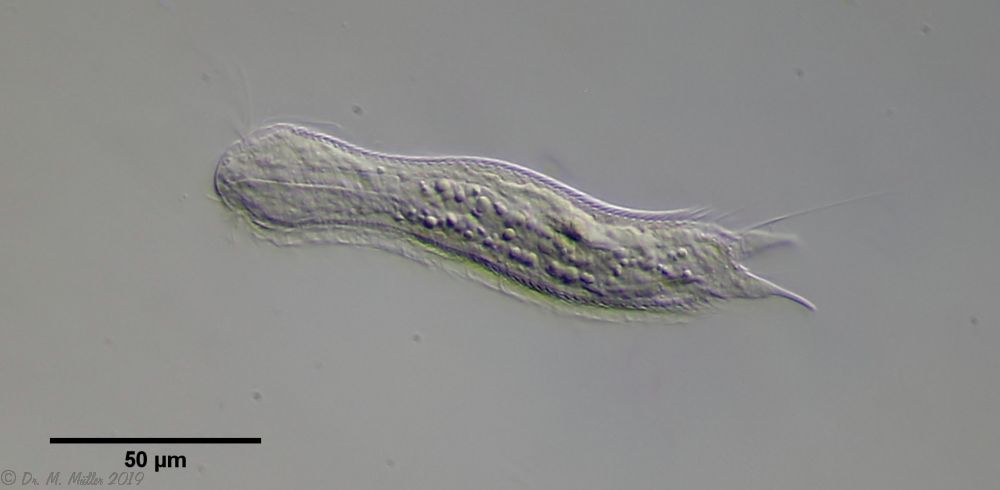
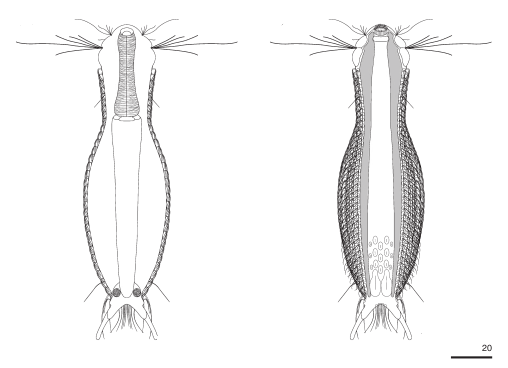
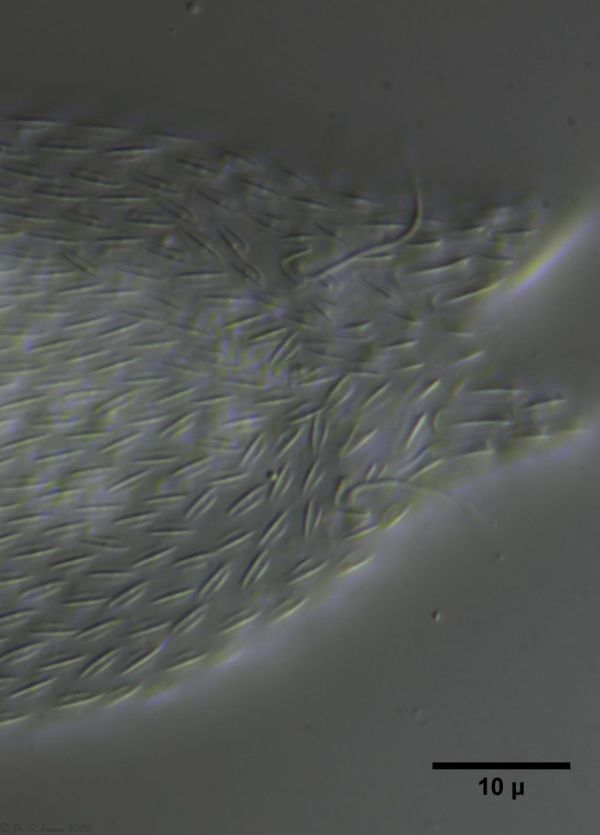
The genus Aspidiophorus is very easily recognized microscopically by the apparent “double contour” of the body. This double contour is produced by the typical style scales of this genus, which consist of a base plate that sits directly on the cuticle. From this base plate rises a thin peduncle, which carries at its end a - usually larger - un-spined terminal plate:
typical scales of an Aspidiophorus, here A. squamulosus.
This results in the typical double outer contour of the animals:
Diagnosis
Body bottle shaped
Furka normal
Peduncle scales
Subsections of Aspidiophorus
Aspidiophorus aster
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Aspidiophorus Voigt, 1903
Species: aster
Length ( bottle ):
Width:
Width of the head ( five-lobed ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( cylindrical ):
Diameter of the mouth ( around ): unknown
Unfortunately, I have no data available for this species!
If you have access to the following literature:
Martin,L.V. 1981. Gastrotrichs found in Surrey. Microscopy 34: 286-300.
I would be very happy if you would send me a copy by Mail!
Unfortunately, I have no data available for this species!
If you have access to the following literature:
Grosso,L.E. 1973. Notas sobre Gastrotrichos Argentinos II. Neotropica 19(59): 87-89.
Grosso,L.E. 1973. Notas sobre Gastrotrichos Argentinos I. Chaetonotus (Zonochaeta) guruguetoi sp. nov. y Aspidiophorus brahmsi sp. nov. Physis 32B: 133-137.
I would be very happy if you would send me a copy by Mail!
Width of the head ( three lobed ): 17 µm - 18.5 µm
µm
Length of the furca: 20 µm
Adhessive tubes: 50% of furca
Pharyx: 32 µm - 36 µm
Diameter of the mouth ( around ): 5.5 - 8.5 µm
Dorsal scales: rhombic petiole scales (3µm long), 41-42 per row, last row in middle with spines (9-14µm), laterally at base of toes a pair of long spines (16-18µm), sometimes with another pair of spines or elongated scales
Ventral scales: Interciliary field with a pair of terminal spines; very many minute keels
Unfortunately, I have no data available for this species!
If you have access to the following literature:
Grosso,L.E. and Drahg,F. 1983. Gastrotricos dulceacuicolas de la provincia de Tucman. I. Chaetonotus soberanus sp. nov. y Aspidiophorus lilloensis sp. nov. Neotropica 29: 189-193.
I would be very happy if you would send me a copy by Mail!
Pharyx ( cylindrical, with small terminal swellings ): 30 µm - 35 µm
Diameter of the mouth ( around ): 3 µm
Dorsal scales: 9 rows of 16-17 peduncle scales each; end plates (6-7 x 2-3 µm) rounded anteriorly, incised posteriorly, no keels; last 2-3 rows terminate in 20 µm spines; lateral on toes a pair of spines (25 µm).
Ventral scales: Ventral intercilliary field with 25 rows of tiny petiole scales
Oecology: Mud dweller
Similar species: A. longichaetus
: Head rather trilobate, no median spines
Dorsal scales: 41-44 rows of 48-50 small peduncle scales each; end plates elliptical, acuminate with keel, overlapping, 3 x 1.5 µm; two 5x2 µm peduncle scales on furca inner side.
Ventral scales: Ventral innercilliary field with two terminal plates (7x4 µm) and 10 rows of narrow, distally pointed keel scales (4x0.75µm)
Dorsal scales: 25 rows of 40-45 rhombic petiolar scales each with lateral and median keels (4x6 µm); petioles 6-7 µm; terminal plates of last terminal rows often enlarged (12-17 µm); posterior end free; between toes 4 short spines on oval scales.
Ventral scales: Ventral innerciliary field covered with petiole scales
Oecology: Sludge dweller
Similar species: distinctive, variable type
Particularities: 3 teeth, eggs smooth (125x74 µm)
Aspidiophorus paradoxus is the largest Aspidiophorus species with about 300µm length.
Dorsal scales
The entire animal is covered with relatively large rhombic peduncle scales.
Cross section
In cross-section, the structure of the petiole scales is clearly visible: the scales sit with a small base plate on the cuticle of the animals, from which rises a thin, hollow pedicel. At the end of the peduncle sits a rhombic terminal plate, with a central keel. At the posterior end of the animal, the terminal plates of the last row of scales are enlarged.
The pharynx of the animal is terminally swollen, and the head is weakly five-lobed with two separate pairs of palpal tufts.
Central view
Ventrally, the strong hypostomion behind the mouth opening is striking. The two ciliated bands split at the head, but the inner branches do not unite in the population I examined. The base of the toes does not bear scales, the adhesive tubes
measure about 50% to 70% of the toe length and taper to a point.
Let’s take a closer look at the scales:
Backscales
In the scales on the back, the rhombic shape of the end plates is most clearly recognizable. Less conspicuous - but typical for the species - is the central keel of the scales.
Cross section shed
In cross-section, the complex geometry of the stem scales becomes clear - base plate, stem and end plate form a very flexible and stable armor. The additional cavity under the outer scales acts like a “crumple zone” and further increases the protective effect.
Cross section scale stems
The stems of the scales consist of hollow tubes that ensure maximum stability with minimum material input - a fascinating example of evolutionary optimization.
The abdomen of the animals is not completely covered with petiolar scales. Rather, they end in the anal region and are joined by simple small, rounded keel scales that are not an obstruction to feces.
Scaling of the abdomen.
At the base of the furca some (according to literature 4) spines protrude into the toe.
The head is almost completely covered with slightly smaller peduncle scales:
Headshed
Cephalion and pleurae are quite small and inconspicuous.
According to literature A. paradoxus has three teeth in the pharynx:
Mouth armament
In the animals I examined, there was only one curved stylet brace, the tip of which protrudes into the lumen of the pharynx and is probably used to open algal cells that are conveyed past it. It is possible that the literature reference to “three teeth” is merely based on a microscopic artifact, as the entire clasp may not be in the focal plane as a whole.
Dorsal scales: 45-53 rows of 29-32 petiole scales each (5-7 µm); no keel, not rhombic, similar shape to that of
P. rhomboides
; Toe base dorsal with keels
Ventral scales: 6 keeled terminal plates; at the posterior end 10-12 rows of keels (4-4.5 µm), otherwise naked
Oecology: between roots; Brazil
Particularities: very small pharynx; body very broad
Dorsal scales: 18-20 rows of 42-45 small petiolar scales each; petioles 1.5 µm; terminal plates 2-3.5 µm long, proximally strengthened margin, distally acuminate, weak median keel; no spines
Ventral scales: Ventral intercilliary field naked, except for a few elongate peduncle scales; two narrow terminal plates(6.5-11.5 µm)
Oecology: Bog, mud
Similar species: delimited by naked ventral intercilliary field
Particularities: without pseudocells; naked ventral intercilliary field .
I have found this belly-harper only once so far in a bog (Sima bog). By the naked ventral intermediate field with long terminal plates the species is well to delimitate:
The round anterior margins of the end plates of the lateral peduncle scales are also well visible. The found animal is with ca. 130 µm a little smaller than the literature specification and all other measurements scale accordingly. The mouth ring is very small and strongly marked. A hypostomium does not exist.
In the median optical section the dumbbell-shaped pharynx can be seen well. The scales are distally long extended (unfortunately the shape of the end plates could not be seen). The granular structure of the two adhesive glands is also interesting.
Width of the head ( five-lobed, pointed cephalion ): ? µm
µm
Length of the furca: 16 µm
Adhessive tubes (tapered, claws): 50% of furca
Pharyx ( cylindrical with slight swellings ): 37 µm
Diameter of the mouth ( around ): 7 µm
Dorsal scales: 53 petiole scales (3 µm) per row; 3 pairs of spines at outer base of toes (20-22 µm).
Ventral scales: Ventral intercilliary field very many petiole scales; a pair of terminal spines (8-10 µm) and many shorter spines protruding into the toe cutout.
Oecology: Mud dweller
Similar species: well demarcated by claws and spines
Dorsal scales: 25-30 rows, each with 60-80 minute petiolar scales; terminal plates unkeeled (0.5-2 µm), elongate, pointed distally; posterior end with elongate keel scales
Ventral scales: Ventral intercilliary field numerous longitudinal rows of minute petiolar scales; two keeled terminal plates (6-7 µm).
Dorsal scales: 12-15 rows of 40-42 petiolar scales each with rhombic terminal plates without median keel (3-4 x 5-6 µm); scale shell ends in anal region; posterior end and toes naked; 3 pairs of short spines at base of toes (8-10 µm).
Ventral scales: 2 terminal keels (5-6 µm), 12 rows of keels without base plates
Oecology: Mud dweller
Similar species: A. paradoxus
: unkeeled basal plates; spination at the posterior end; mouth grafting
Of the (acc. (Schwank, 1990))
twenty limnic Aspidiophorus species only four are listed in the “official” species lists for Germany. Another species was described some time ago by Michael Plewka. The species Aspidiophorus squamulosus has been found so far only in Poland and in western France - so it is no wonder that this animal is also native to us, between the previous finding areas.
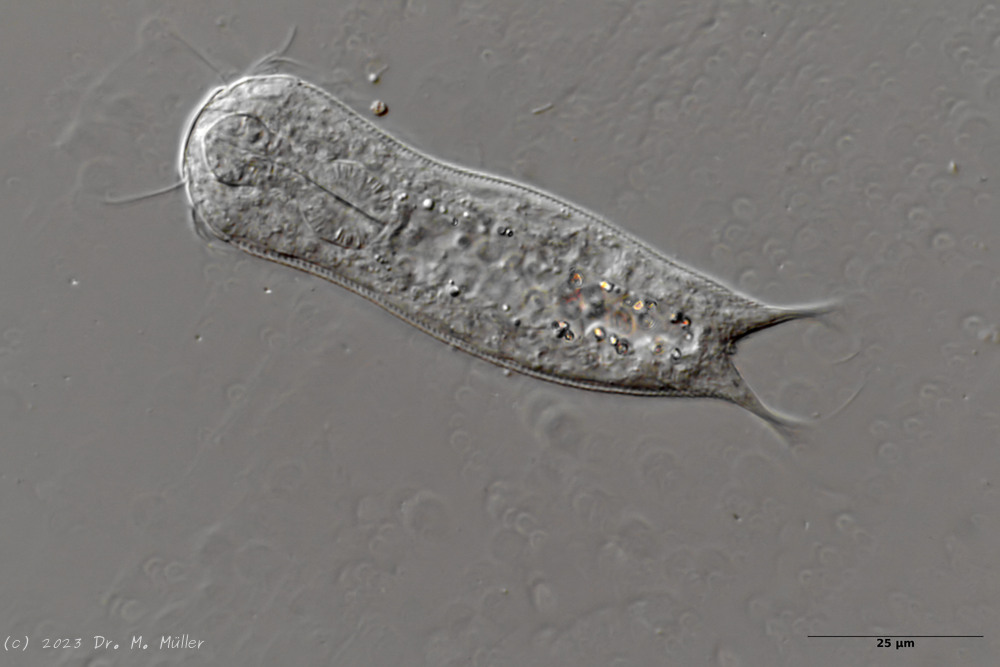
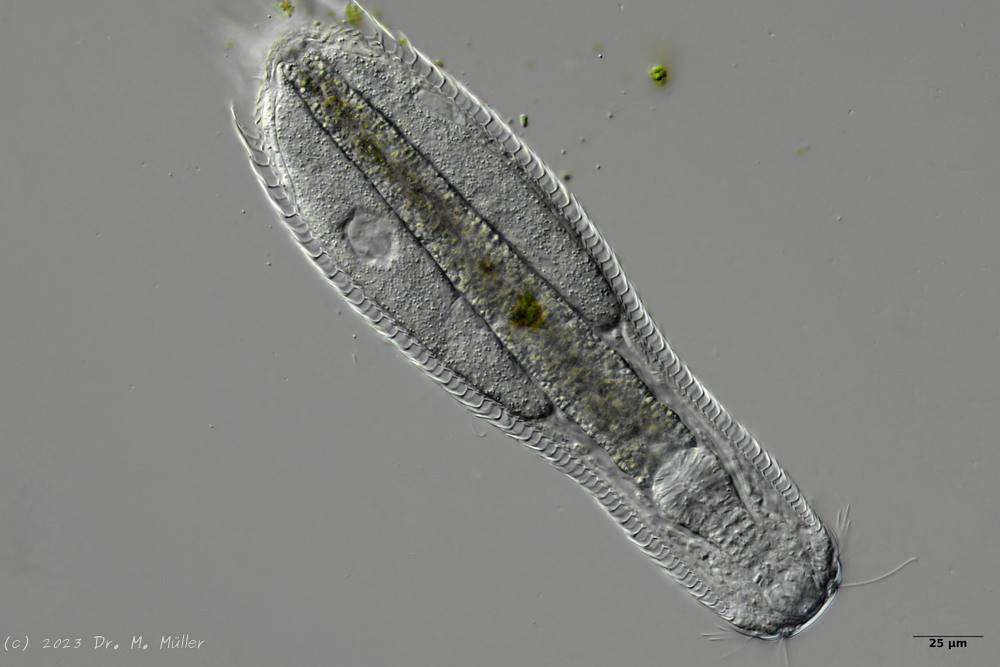
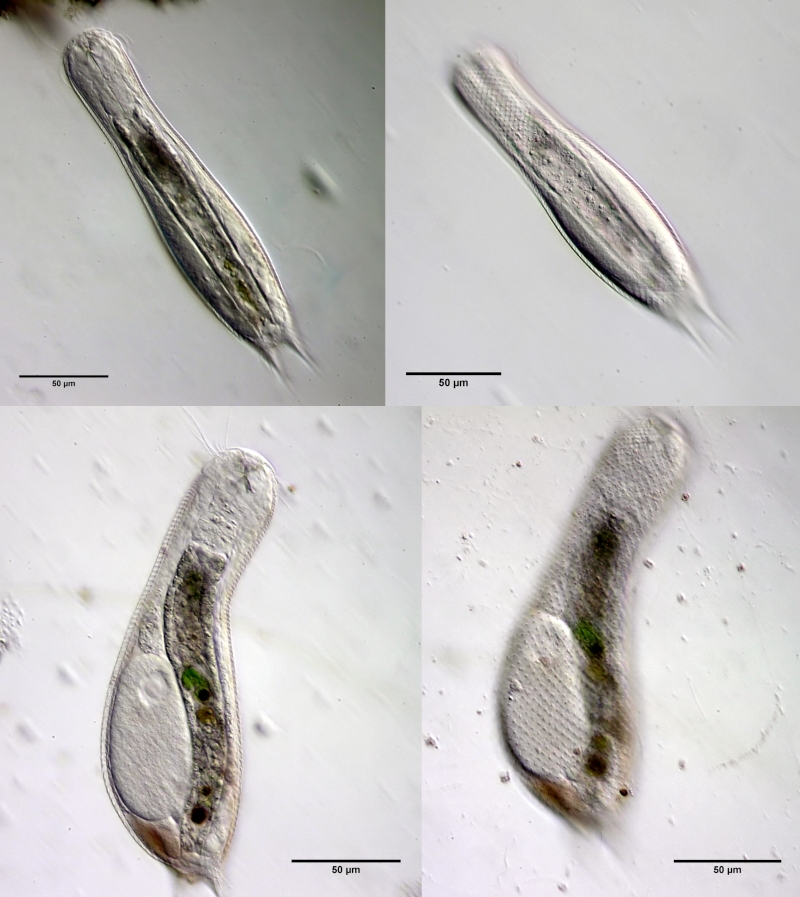
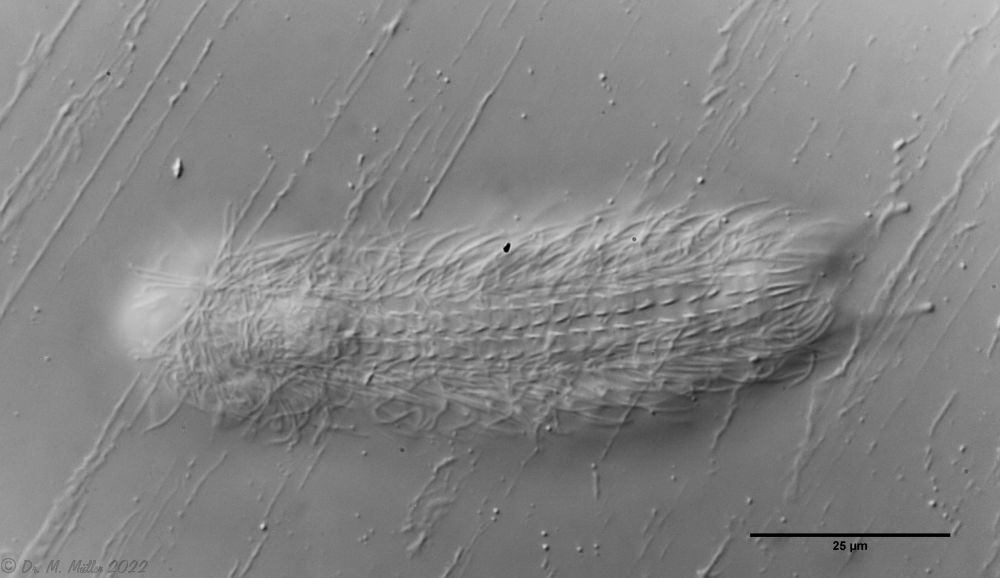
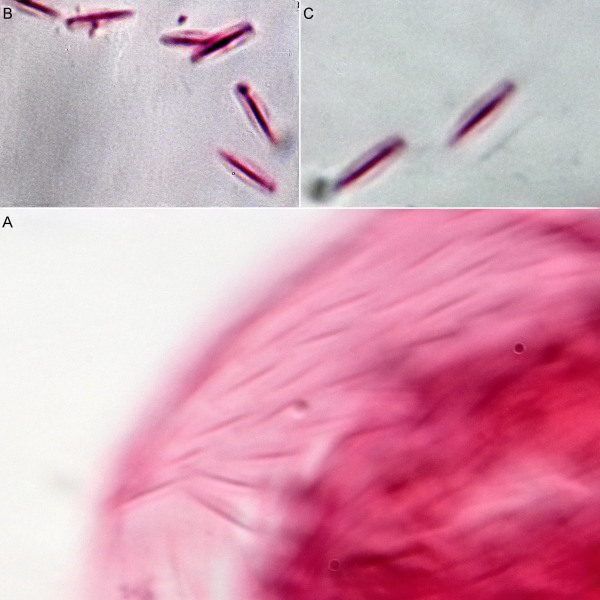
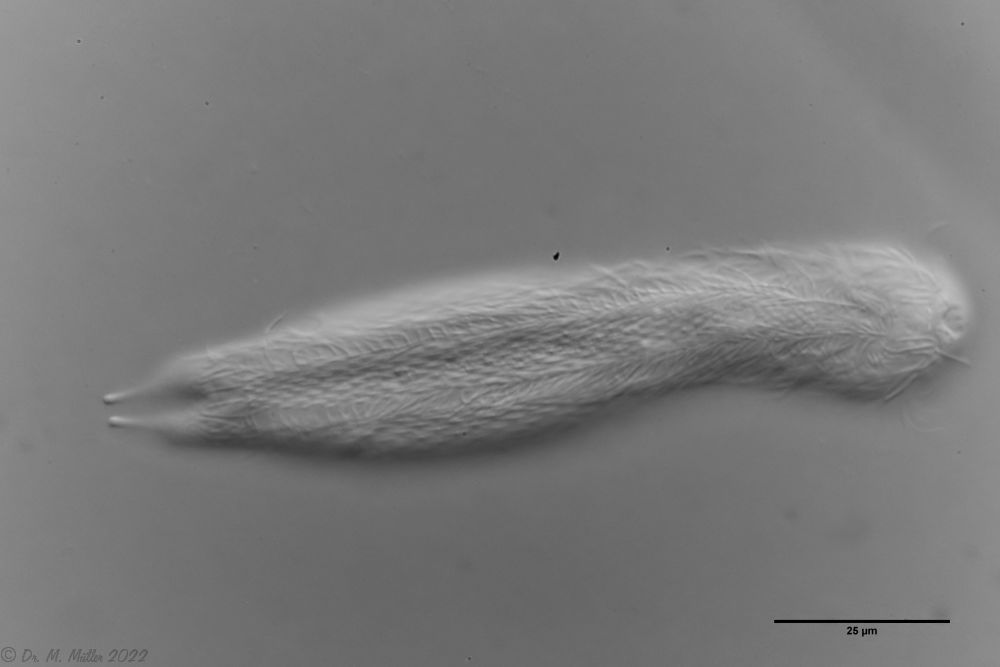
Fig. 1: Aspidiophorus squamulosus; optical section and scale image.
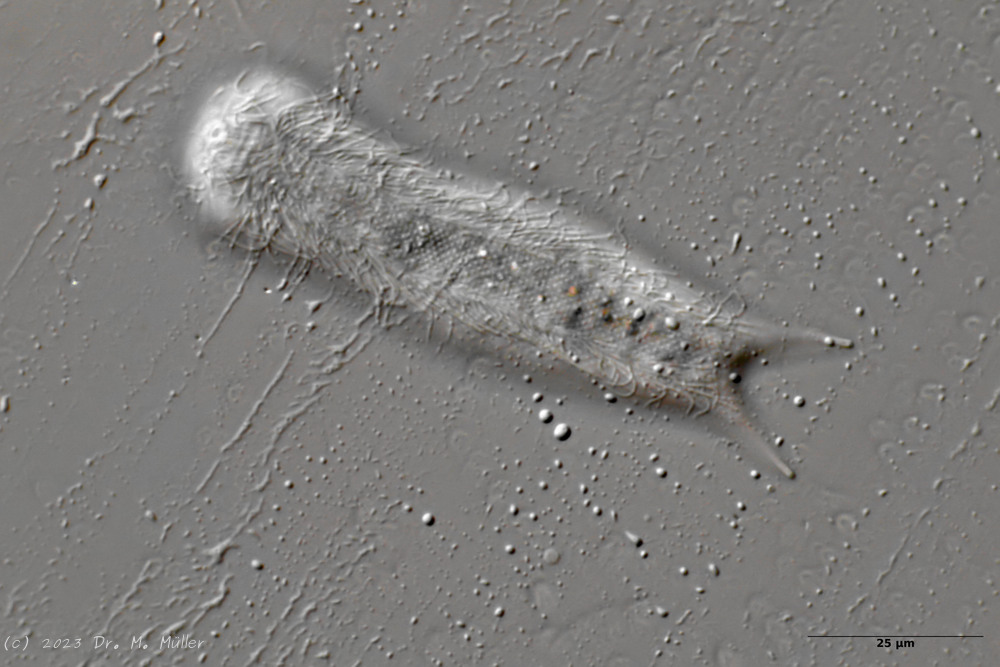
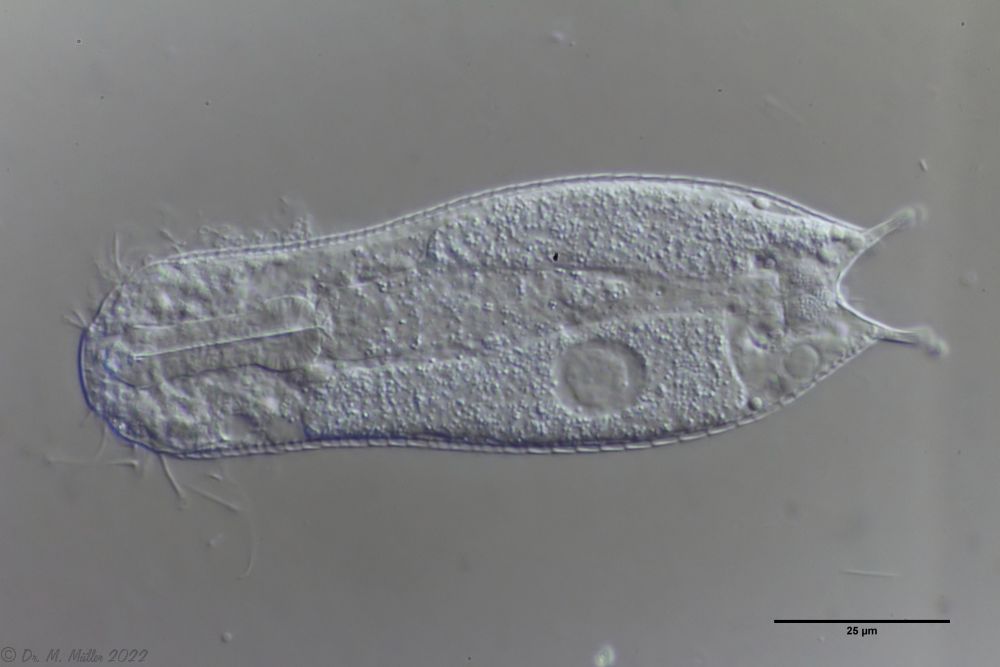
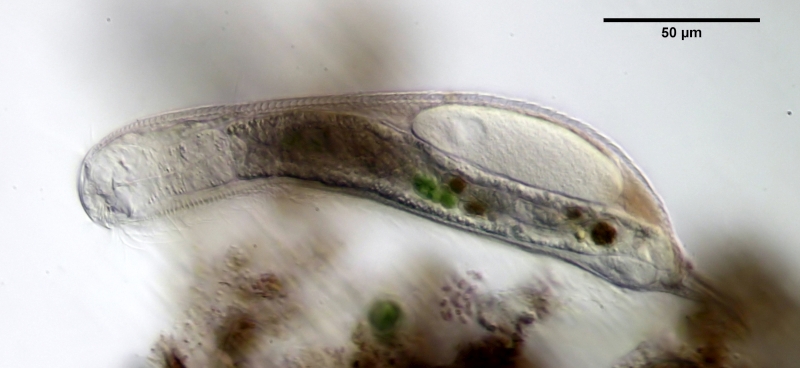
The internal structure of the animal becomes quite clear in the side view:
Fig. 2: A. squamulosus, side view.
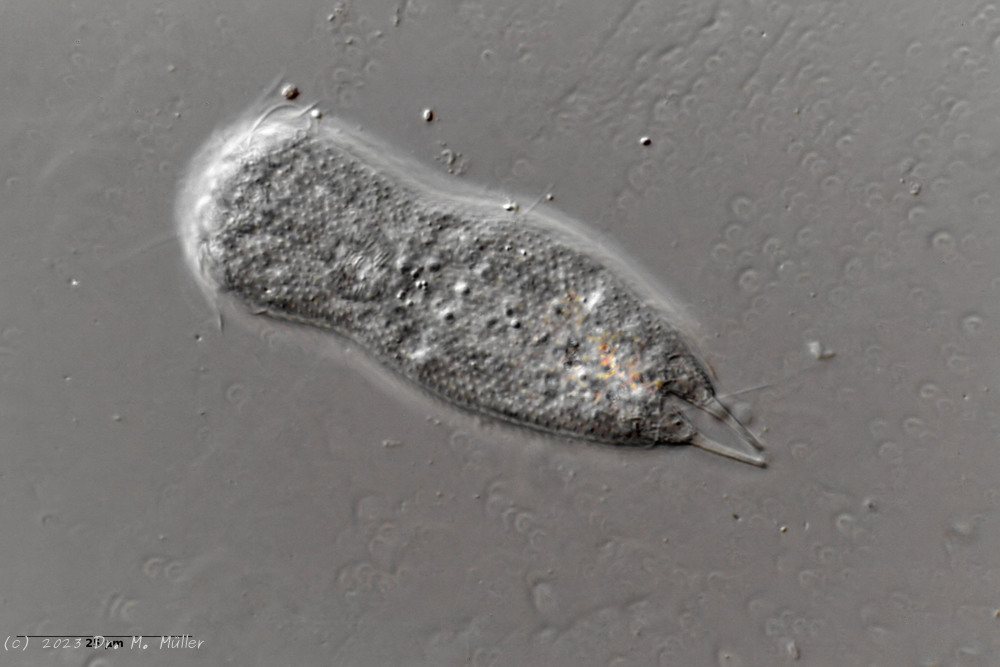
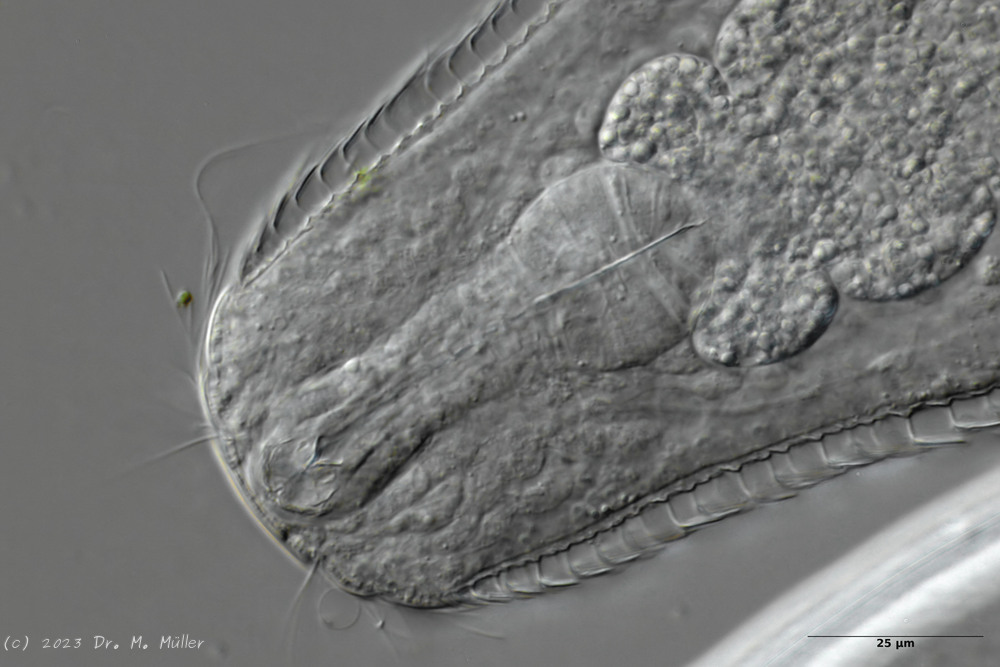
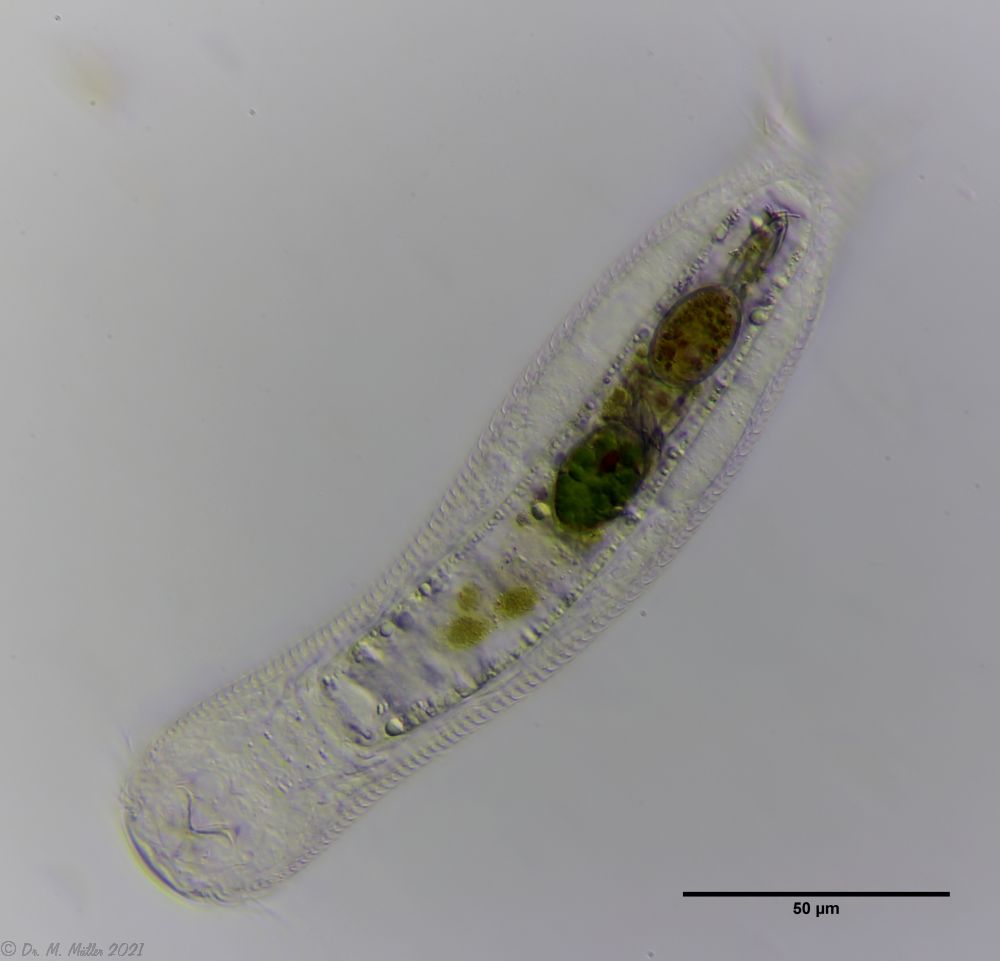
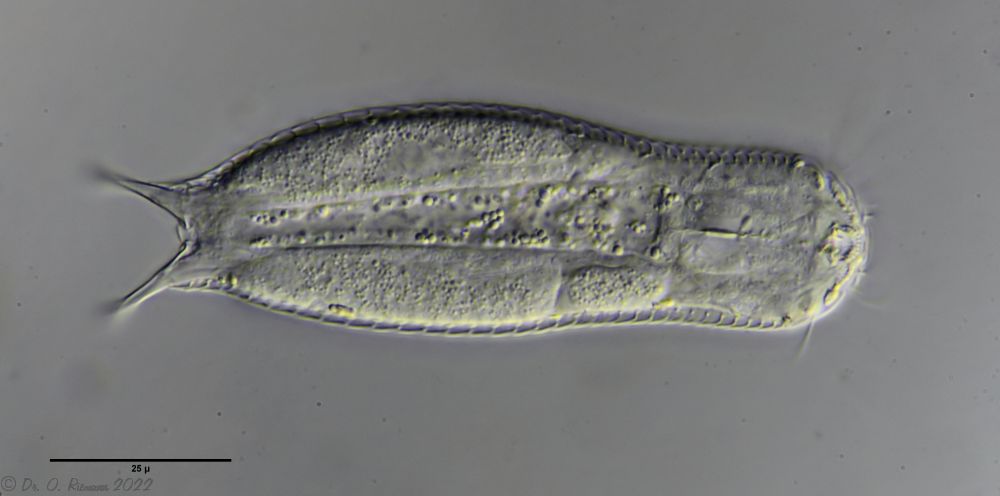
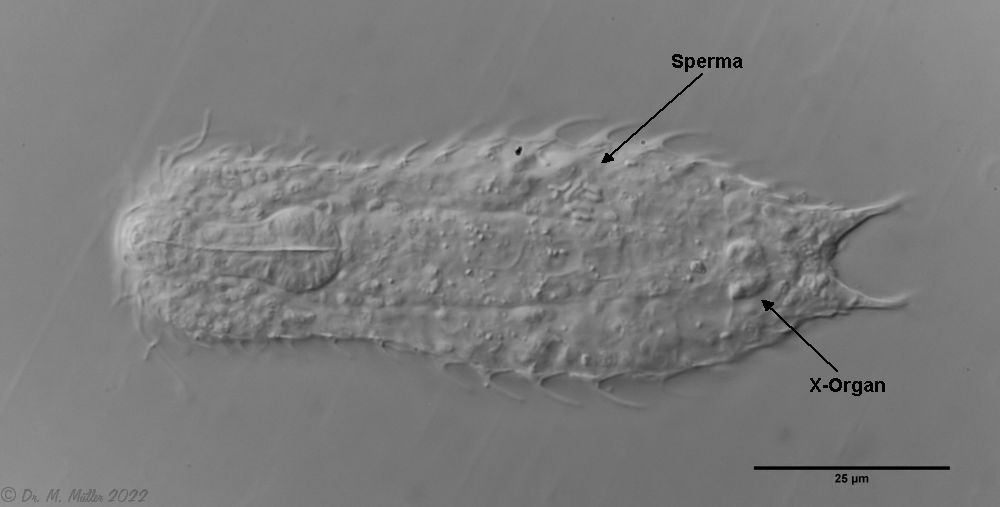
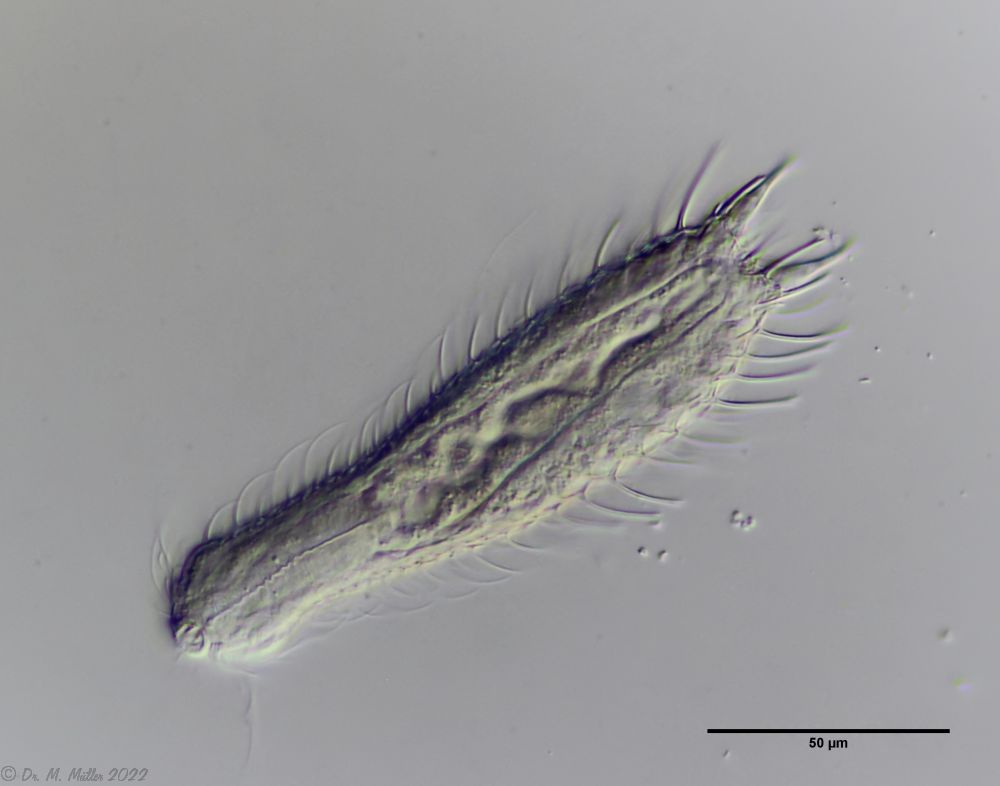
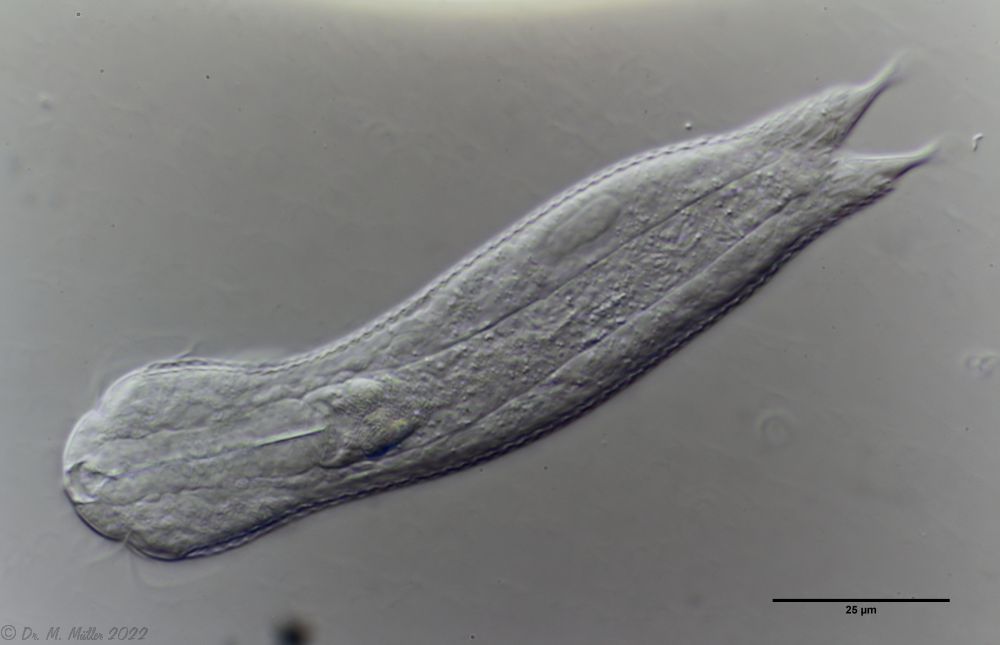
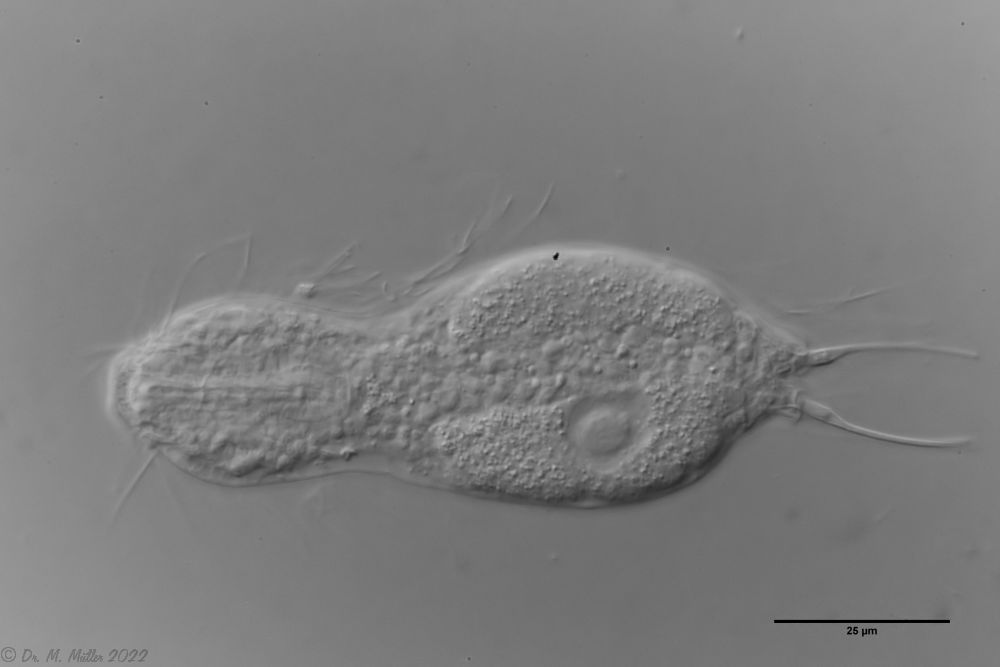
A. squamulosus is with ca. 230µm somewhat smaller than the similar species A. paradoxus and is clearly distinguished by the three spines each above the toes (pic. 3, ob. li. and mi. r.). In addition, the terminal plates of the petiolar scales do not possess a median ridge. Also the prominent, furrowed hypostomium (Fig. 3, ob. r.) is typical for this species. Some animals I found were in their “hermaphroditic phase” (post-parthenogenetic phase) and showed a very large X-organ and sperm bundle (Fig. 3, mi. l.). One animal also carried a very large egg (with a pronounced nucleolus), which it laid during the approximately 10-day observation period. Unfortunately, I was not able to observe the oviposition itself (also the egg was not found in the specimen).
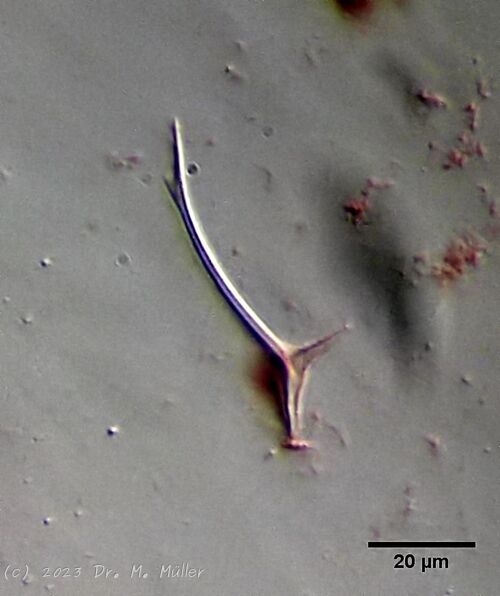
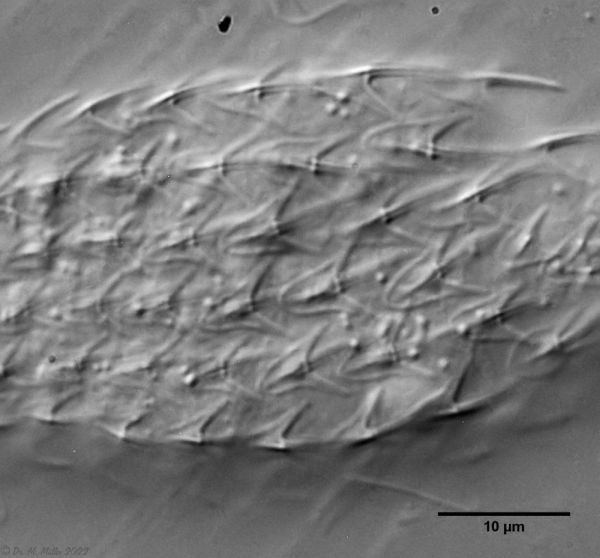
Figure 3: A. squamulosus: details;
up. l.: toe spines; up. r.: hypostomium
mi. l.: X-organ and spermathecae (marking); mi. r.: toe spines and toe scales;
low. l.: transverse section of scale carapace; low. r.: stylet (mark)_
As is often the case, there were some deviations from the species description in the literature. With up to 50 generations per year and years of isolation in a small garden pond, I find minor deviations not surprising. However, it is possible that some details were not noticed in the rare, earlier observations. For example, the base of the toes should be unscaled (Fig. 3, mi. r.). However, the specimens I observed show many very small, roundish and close-scaled scales. Only the conspicuous peduncle scales do not appear at the posterior end of the animal.
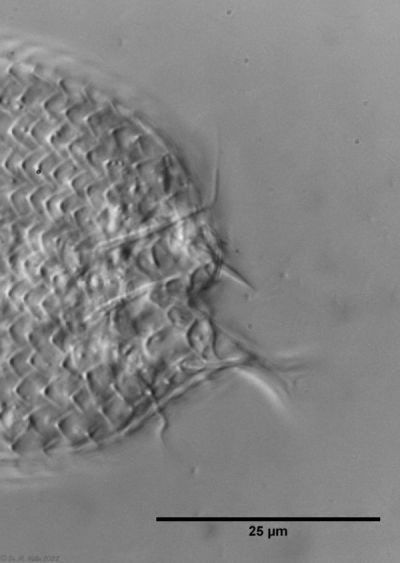
In some Aspidiophorus species movable “teeth” in the mouth tube are described. Although A. squamulosus should be toothless, two actively movable styletes could be observed, the tip of which ended in the mouth tube and reminded of tardigrades (Fig. 3, un. r.).
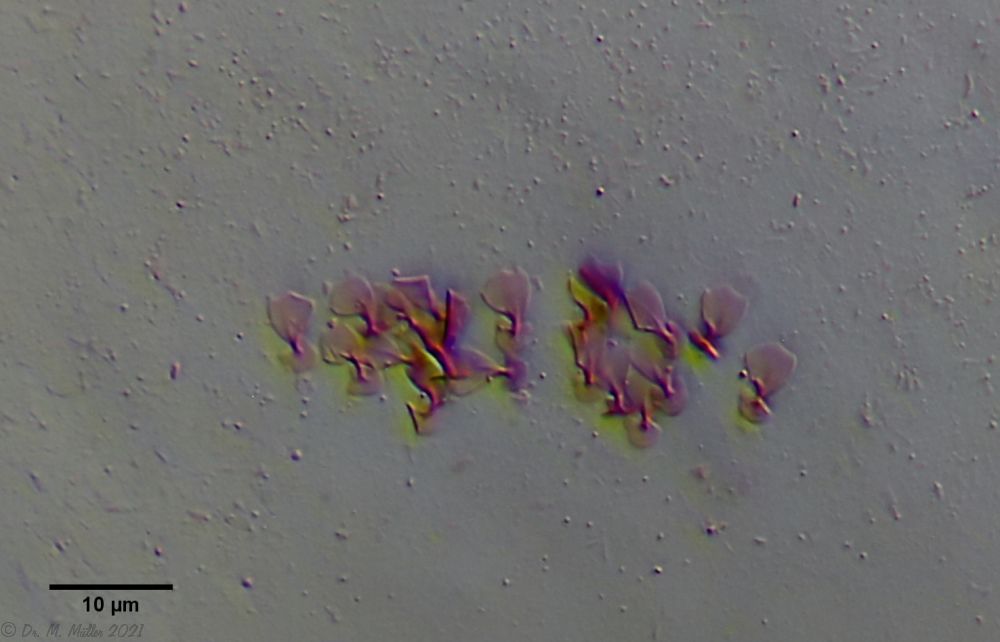
The peculiar stipe scales of the genus Aspidiophorus are not to be grasped in their three-dimensional beauty in the scale association. Therefore, I subjected a pitiable specimen of A. squamulosus to scale analysis:
Fig. 4: A. squamulosus; colored and - unfortunately - flattened single scales.
Since during a scale analysis just the very small scales are pressed rather flat, I tried a drawing reconstruction of the three-dimensional structure:
Fig. 5: A. squamulosus; bottom: composite scale image; top left: isolated and stained scales; top right: drawing reconstruction of a single peduncle scale.
Another point that has remained open so far is the diet of A. squamulosus. The main diet consists of relatively large unicellular algae or eye flagellates. To swallow this large prey, the mouth of the animals is equipped with movable lamellae that can be folded outward for feeding (cf. Captochaetus). This allows the diameter of the mouth opening to be greatly expanded, unlike most gastrotrichs. During the swallowing movement, the stylet is pressed into the mouth tube and the prey cells are guided past it. In the process, the hard cell wall is probably perforated so that the digestive secretions in the intestine can penetrate the cells.
Fig. 6: A. squamulosus; intestine filled with ocular flagellates.
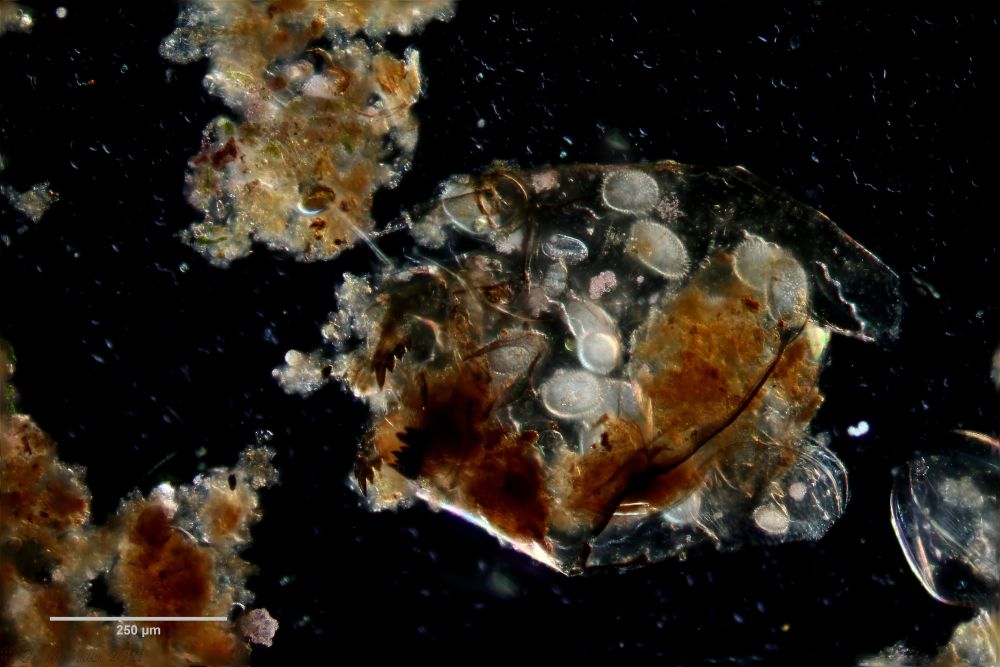
If one examines the mud of stagnant waters microscopically, one always finds gastrotrich clutches in empty shells of water fleas with sometimes dozens of eggs.
Fig. 7: A. squamulosus; focal sac of a gastrotrich clutch containing eggs of at least three different species.
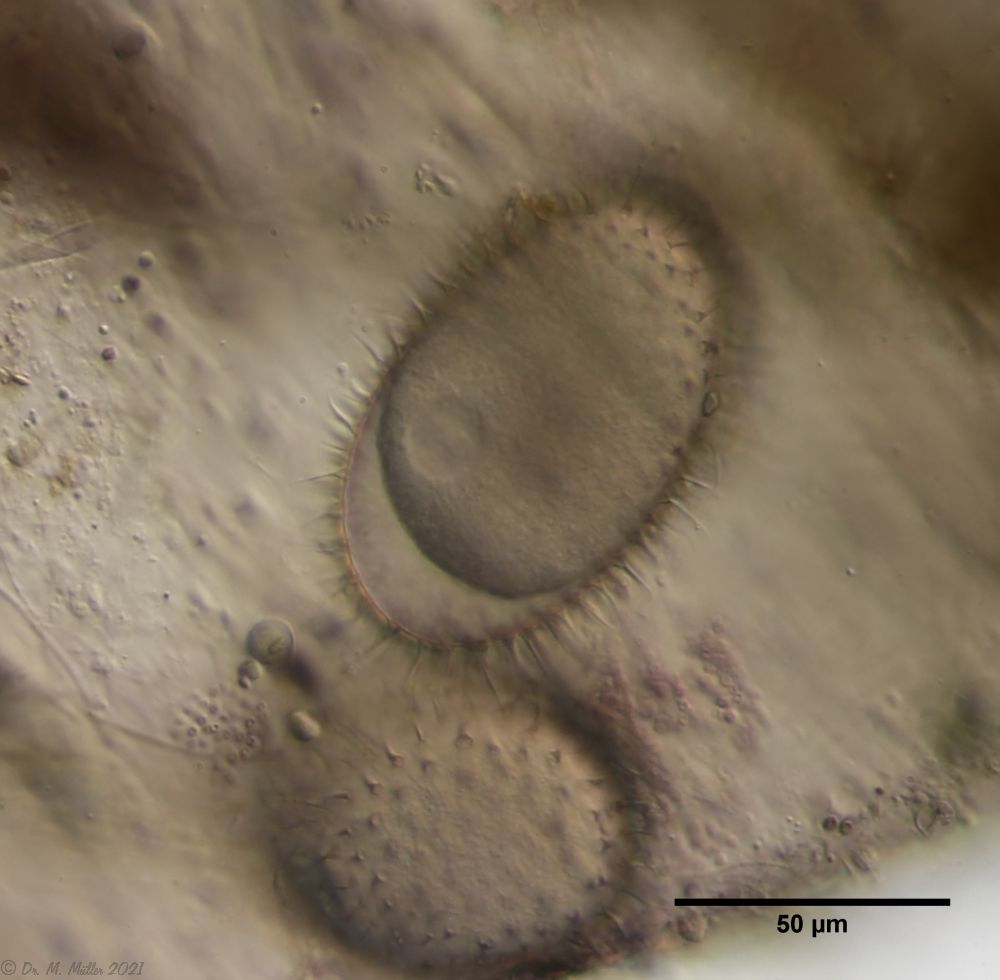
Often these clutches consist of eggs of different species. Apparently, a number of species prefer to lay their eggs in water flea shells where other gastrotrichs have already laid their eggs. In such a clutch often relatively large (95µm x 55 µm), spiny eggs are noticeable, which have not yet started embryonic development and probably represent permanent eggs.
Fig. 5: A. squamulosus; permanent egg.
One egg of the shown clutch started to develop and turned out to be an egg of the species A. sqamulosus. Therefore I can show here the eggs belonging to the species.
Dorsal scales: 15-20 rows of 38-40 very small peduncle scales each; endplates very delicate and unkeeled (2.5-3 µm); strongly bent peduncles -> scales protrude like bristles
Ventral scales: Ventral intercilliary field 9-10 rows keels; anteriorly mostly naked; no terminal plates.
Oecology: creek
Similar species: well demarcated by protruding scales
Particularities: protruding scales; body cylindrical
Dorsal scales: 15-17 rows of 33-40 petiole scales each; end plates unkeeled 2-5µm, rounded distally and proximally; base of toes naked; 2 pairs of thick spines (8-10µm and 4-5µm) on oval scales with two keels.
Ventral scales: 2 terminal keels (7.5-11µm); in gut region 8-9 rows of tiny keels, otherwise naked
Oecology: montane lake
Similar species: delimitable by spines
Particularities: 2 pairs of spines at the rear end
A. tetrachaetus is a very rare gastrotrich, so far confirmed only for Poland.
A. tetrachaetus: dorsal ((c) Ole Riemann)
The dorsum of the animals is covered with relatively large, unkeeled and rounded pedunculate scales, which do not continue to the base of the toes at the posterior end. A pair of large spines is conspicuous at the posterior end of the scale coat.
A. tetrachaetus: t.s. ( Ole Riemann)
In the cross section the structure of the petiole scales can be seen well.
A. tetrachaetus: ventral (copyright: Ole Riemann)
The ventral intermediate field of the animals is covered only in the intestinal region by 8-9 longitudinal rows of small keels. The pharyngeal region is naked. Two terminal keels form the posterior shoot of the ventral scaling.
A. tetrachaetus: two pairs of spines at the hind end.
Only a closer look at the dorsal toe base of the animals reveals two additional, smaller spines.
A. tetrachaetus: single tooth in oral cavity.
Contrary to the description in the literature, a single tooth can be seen protruding asymmetrically from the oral cavity in the animals at Sima Moor.
Adhessive tubes (three-part (17, 10, 7 µm)): 80% of furca
Pharyx ( dumbbell-shaped ):
Diameter of the mouth ( around ): unknown
Dorsal scales: 7-9 rows of 17-18 unspined scales each; anterior 2/3 round to hexagonal, last 5 rows 3-5 larger scales each.
Ventral scales: Hypostomium large, Ventral intercilliary field with 3-5 rows of 15-17 spineless, rectangular scales each; 2 large keeled terminal plates followed by a pair of narrow, long keels.
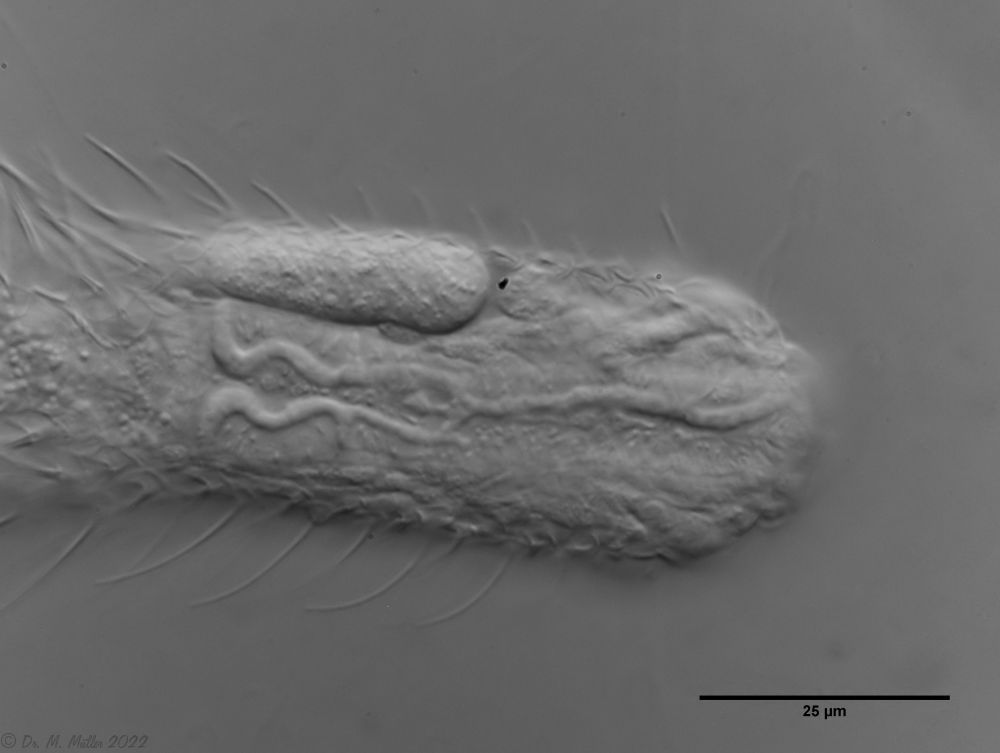
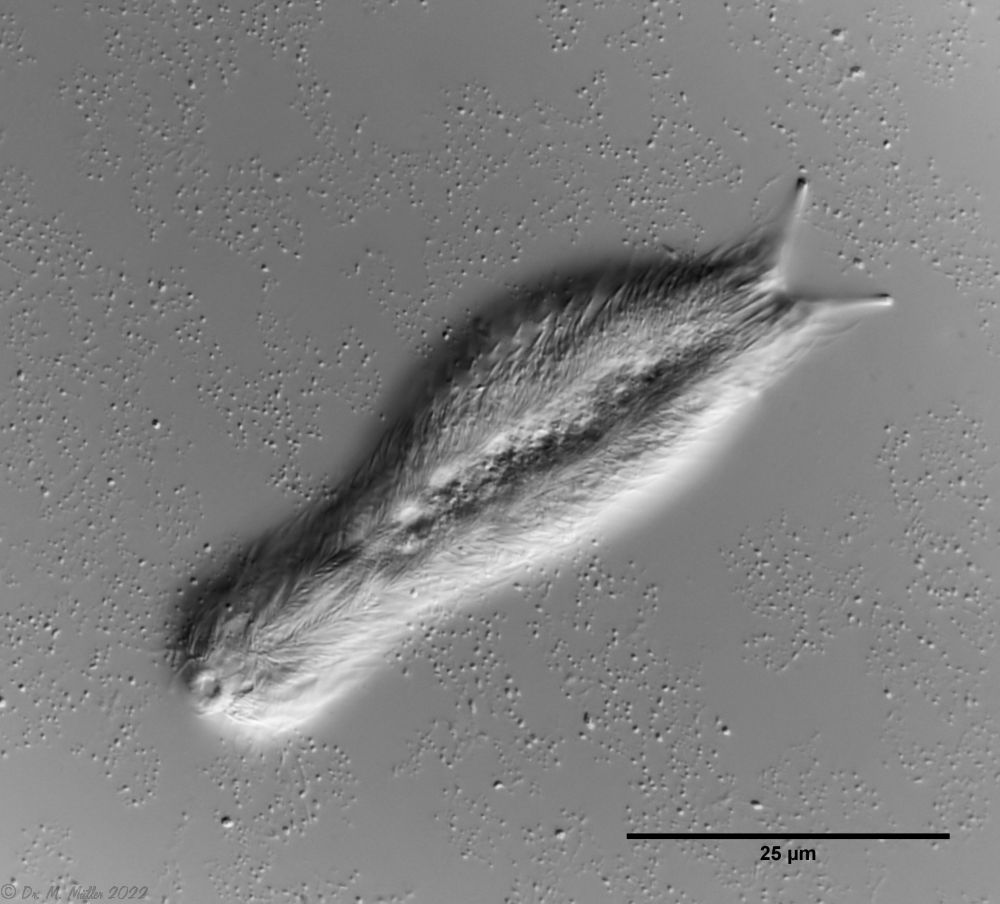
Dorsal scales: Head: 18-19 rows; body 10-11 rows with a total of 33-40 thin, strongly overlapping heptagonal to rectangular scales (19µm x 9µm), each bearing a spine (2µm - 40µm) with a secondary tip; short spiny field at the rear end
Ventral scales: ventral intermediate field with two curved, keeled terminal plates;8-9 rows of keel scales (partly spiny); oval, smooth scales in the pharynx region
Particularities: Distinct system of pharyngeal veins
Chaetonotus hoanicus is a very rare gastrotrich that has so far only been observed by Schwank. I would therefore like to describe it in more detail. The main characteristic of Ch. hoanicus is the abrupt transition from an unspined (or shortly spined) head area to a relatively long spined back:
Ch. hoanicus: Change of the dorsal spination
The scaling of the neck/head area consists of almost round, strongly overlapping scales with only an extremely short - often only perceptible - spine:
Ch. hoanicus: Change in the dorsal scales on the neck
With an abrupt transition to long spiny scales the area of the trunk scales is clearly separated. The trunk scales are relatively large, roughly pentagonal and have a large cut-out at the rear end. Spines approx. 15-20 µm long arise from its tip, which are provided with a secondary tip distally.
Ch. hoanicus: Modification of the dorsal trunk spination
In the anal region there is a short spined field. Some conspicuous, longer spines protrude into the toe cutout.
Ch. hoanicus: Change in the dorsal spination of the anal region
The exact scale shape only becomes clear when it is possible to detach scales:
Ch. hoanicus: Individual dorsal scales from the dorsal region
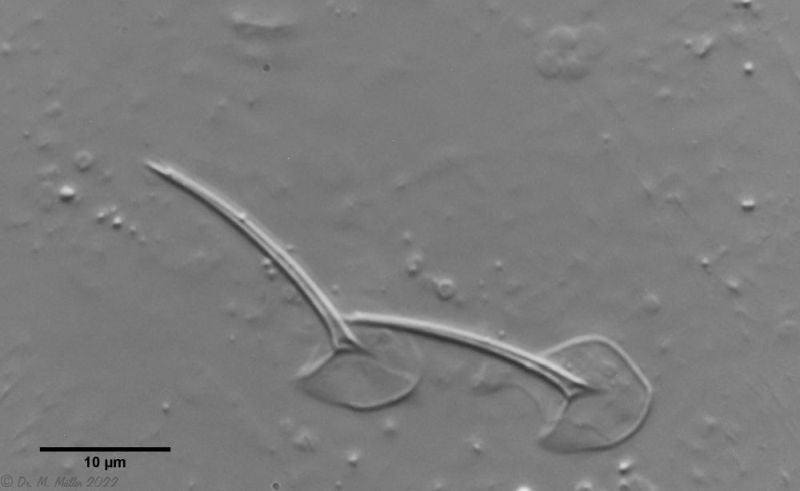
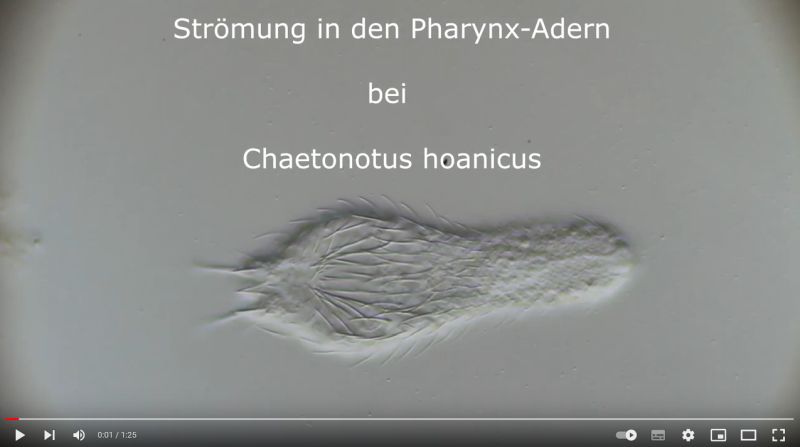
The particularly pronounced tuning fork-shaped structure of the pharyngeal surface, which connects the oral cavity with the glands in the pharyngeal wall (see Pharynx), is conspicuous:
Ch. hoanicus: Structure on pharyngeal surface
If one films a living specimen of Ch. hoanicus with the focus on these “veins”, a clear flow within these structures becomes visible, indicating a gland-like function:
Dorsal scales: 17-19 rows of 38-40 circular scales each with distal cutout and short and simple spines
Ventral scales: 2-3 pairs of elongated, keeled terminal plates; 5-7 rows of small, round keel scales
Oecology: very europhilous species; standing and flowing waters; moor; common
Similar species: well-defined due to stocky shape and bare toes
Particularities: Bare toes; insufficiently described
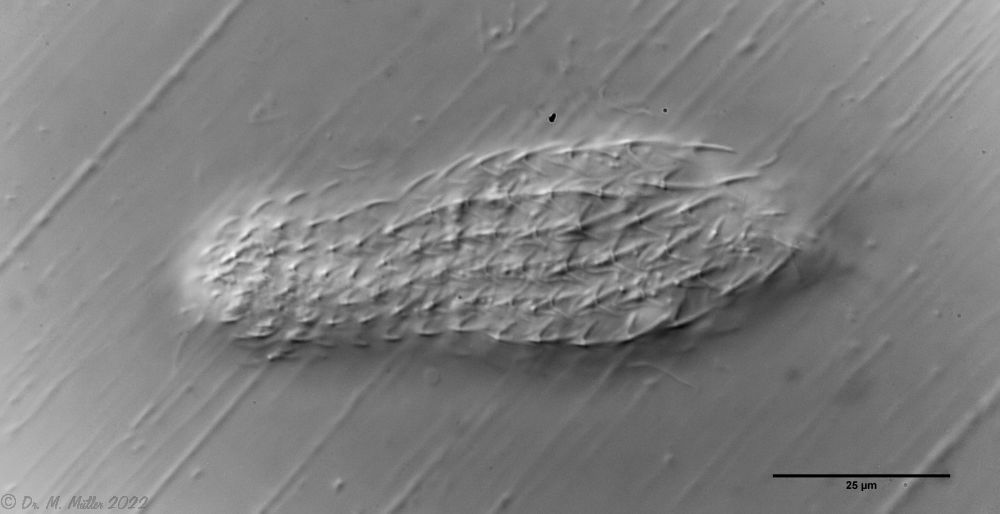
The small and common gastrotrich Chaetonotus (Ch.) multispinosus is characterized by its short-spined, roundish scales, which are cut out at the posterior end:
Ch multispinosus: ventral
The short spines that give the species its name can be seen in cross-section. The short, club-shaped pharynx is very typical:
Ch multispinosus: cross section
The ventral intermediate field is covered with a few rows of roundish, keeled scales. The posterior end is formed by two larger, short spiny plates, which are bordered by two pairs of small, also short spiny keel scales:
For the subgenus Hystricochaetonotus are the relatively large, non-overlapping scales with a long spine. This subgenus owes its name to this spiny appearance (hystix = porcupine).
The scales are in typical arrangement. Their shape is trilobate with a central keel, at the end of which arises a long spine - usually with a secondary tip:
typical three-lobed scales in Hystricochaetonotus - here
C. (H. ) persetosus
Dorsal scales: 13 rows of 13-14 three-lobed scales (6.5µm x 5µm) with simple spine (3µm - 10µm); typical scale group for Hystricochaetonotus without spines at the posterior end
Ventral scales: ventral intermediate field with two rows of 20 rectangular scales each (1.2µm - 4µm); two keeled terminal plates (length 6.4µm - 6.8µm)
Oecology: Moor
Similar species: all Hystricochaetonotus; precise diagnosis required
Particularities: Spines without secondary tip; unique scaling of the ventral intermediate field
The small gastrotrich Chaetonotus (H.) italicus, previously only known from Italy, is relatively common in the Sima bog.
Ch. italicus: dorsal
Many of the specimens found were in their post-parthenogenetic (hermaphroditic) phase and bore both a distinct, bipartite X-organ and rod-shaped spermatoids. The pronounced dumbbell-shaped pharynx is typical of the species:
Ch. italicus: t.s.
The ventral intermediate field is covered with two rows of rectangular scale plates, which become more indistinct in the neck area. Two keeled terminal plates form the end:
Ch. italicus: ventral
Although the scale spines do not have a secondary tip, the classification in the subgenus Hystricochaetonotus is clear due to the typical scale shape:
Ventral scales: 2 long, thin terminal plates with short spines; 7-8 rows of small keel scales (1µm - 3µm)
Oecology: Moor, bank moss
Particularities: Toes naked; rigidly protruding spines are very typical
The very rare gastrotrich Chaetonotus (H.) murrayi is only known from two localities (in Poland and near Berlin). The strong and quite long two-pointed spines protrude in a spear shape and give the animal a very defensive appearance.
Ch. murrayi: cross section
This is particularly noticeable when focusing on the spines on the back:
Ch. murrayi: dorsal
The approx. 20µm long spines have a lateral secondary spine near the main spine:
Ch. murrayi: focus on the spine tips
The focus on the scales shows the typical three-lobed scale shape of this subgenus. In contrast to the findings in the literature, the entire head of the animals is also covered with long spines:
Ch. murrayi: dorsal scaling of the head / neck area
The spination is also clearly pronounced at the posterior end. Unlike many members of the subgenus Hystrocochaetonotus this species lacks the unspined dorsal keel plates:
Ch. murrayi: dorsal scaling of the hind end
The ventral intermediate field bears 6 - 7 rows of small keel scales and is terminated by two spiny terminal plates.
Ch. murray: Scales of the ventral intermediate field
Dorsal scales: 7 rows on the head, 12 on the trunk, each with 14-15 three-lobed, keeled scales (3-6 µm), which do not overlap and are short and delicately spined (10 µm); two rows of longer spines at the head end. Very long spines with two secondary tips (26-50 µm) in a relatively narrow field on the anterior trunk
Ventral scales: 2 elongate-oval, keeled terminal plates (6.6µm x 4µm) with two simple spines; 4-6 rows with 40 oval keel scales each. Own observation: in pharyngeal region rectangular transverse claspers
Oecology: Moor, plant-rich small bodies of water
Similar species: C. (H. ) anomalus
is possibly synonymous; differs only by the absence of the two scale rows with longer spines on the head (which was present in my observations) and by scale claspers below the pharynx (which are also present in Ch. novenarius in my observations).)
Particularities: easily delimited by the long spines that emerge from a narrow field
Chaetonotus (H.) novenarius is immediately recognizable by its nine very long large spines.
Ch. novenarius: dorsal
The nine large spines each have two secondary tips and extend beyond the end of the animal’s toes. Ch. novenarius is the Chaetonotus with the longest spines relative to the body length (approx. 2/3 of the body length):
Ch. novenarius: dorsal large spines
Although the scales and spines clearly identify the animal as Ch. novenarius, the ventral scales of the found animal are puzzling:
Ch. novenarius: ventral
Striking are the rectangular scale plates in the pharynx area, which Schwank only shows for the very similar species Ch. anomalus. It is possible that this species is a synonym of Ch. novenarius, as (Schwank, 1990)does not rule out.
Dorsal scales: 5 rows of 15-18 transverse oval scales (5 x 10 µm) each, not touching; on head dorsally and laterally 5-7 characteristic spiny scales; small spines 6-7 µm; groups of large spines 8-22 µm.
Ventral scales: Ventral intermediate field on head 3 touching plates; otherwise elliptical plates; in gut region sometimes 3 deer of smaller elliptical plates.
Oecology: Bog waters, rare, very variable
Particularities: very variable with respect to spine size - possibly a number of subspecies
A very rare and strange ventral hardy is the nearly rotund Chaetonotus (P.) acanthocephalus with its irregular spination and large, round scales:
Ch. acanthocephalus: focus on the dorsal scale
Ventrally, the animal bears large cuticular plates on the head that change to round, short-spined scales toward the posterior end:
Ch. acanthocephalus: ventral
If the animal is macerated with glacial acetic acid, the scale shape typical for the subgenus Primochaetus becomes particularly clear:
Dorsal scales: 10 rows of 20-25 strongly overlapping, rounded triangular scales (5 µm - 8.5 µm) with strongly convex margin (margin appears double) with short spines (7 µm); 7 - 12 simple girdle spines (6.5 µm - 16.5 µm) in alternating rows; scales behind spine girdle mostly without spines
Ventral scales: 4 elongated terminal plates with keel; 8-9 rows of small keel scales
Oecology: Moor, on mud
Similar species: well demarcated by spine belt and double scale edge
Particularities: Spiked belt and double scale edge
A further representative of the subgenus Primochaetus is the small abdominal hardy Ch. (P.) acanthodes, which is notable for its distinct, spineless scales with a “double” anterior contour and a belt of long spines on the abdomen:
Ch. acanthodes: dorsal scales
Only in the lateral view one recognizes that the spiny belt covers the entire dorsal side of the animal:
Ch. acanthodes: side view
The ventral intermediate field is covered with approx. 8 rows of small, roundish keel scales. The rear end is formed by four rectangular, narrow keel plates:
Ch. acanthodes: ventral scales
In cross-section, it becomes clear that the scales are strongly curved. This results in the “double contour” in the optical section. The animals I found have one or two teeth at the entrance to the pharynx:
Dorsal scales: 7 rows of fe 20 thin, transverse-oval to obovate scales, only slightly touching each other; spines (10-39 µm) originate slightly behind scale center; secondary tip relatively far from distal end; three pairs of long lateral spines (34-46 µm) overhang toes by twice their length; between toes 3 dorsal spines slightly overhanging toes
Ventral scales: Ventral intermediate field with +11 rows of short spiny tiny scales
Particularities: brood care: decorating the eggs with detritus; var. bernensis has no secondary tips
Ch. cordiformis shows in cross section the typical structure of limnic gastrotrichs. Remarkable are the relatively few, long and strongly curved scale spines:
Ch. cordiformis: cross section
The side view shows the enormous length of the spines which are provided with a secondary tip:
Lateral view(ChCordiformis2.jpg)
Ch. cordiformis: side view
These long spines make it very difficult to take good photos of the animals, because pinning them down with the coverslip strongly disturbs the natural arrangement of the spines. Therefore, an attempt should be made to capture the appearance of the animals free-swimming:
Ch. cordiformis: free swimming specimen with focus on the scale spines
High resolution images are only possible with somewhat pressed specimens. Only here the roundish scale shape with the scale spines provided with a secondary tip becomes visible. The secondary tip sits relatively far away from the spine end of the animals.
Ch. cordiformis: spiny scales with secondary tip.
Many of the specimens found had peculiar spherical bodies inside, which I think are (permanent forms? of) parasites:
Ch. cordiformis: spherical inclusions.
The scales of the animals can be judged well only after they are detached. Here you can recognize the name-giving “heart shape” of the roundish scales with an indentation:
Ch. cordiformis: heart-shaped scales
Ch. cordiformis is one of the few gastrotrichs that performs a kind of “brood care” after egg laying. After oviposition, the mother transports detritus from the vicinity of the oviposition site in her pharynx to the egg and sticks this material to the eggshell. As a result, the eggs are completely covered by detritus and protected by it from enemies:
Ch. cordiformis: brood care - start movie by clicking.
Dorsal scales: 5 Reihen mit je 20 einfachen, dicken und kurzen Stacheln; lateral 12 Großstacheln: 2 Paar am Hals, 1 Paar in der Mitte, 3 Paar am Hinterende
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Schimkewitsch,W. 1886. Uber eine neue Spezies Ichthydium. Nachr. Gesell. Freunde Naturw. Moskau 50: 148-150.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus crinitus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: crinitus
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: unzureichend beschrieben und so nicht wieder zu erkennen!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Sudzuki,M. 1971. The Gastrotricha of Japan which live in the capillary water of the interstitial system: II. Bull. Biogeogr. Soc. Japan 27: 37-41.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus dentatus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: dentatus Tretyakova, 1992
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Chaetonotus elegantulus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: elegantulus Remane, 1936
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: sp. inquirenda; jegliche Abbildung und Beschreibung fehlt!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Remane,A. 1936. Gastrotricha, In: H.G. Bronns, ed. Klassen Ordnungen des Tierreichs, Band 4, Abteilung II, Buch I, Teil 2, Lieferungen 1-2. Akademie Verlagsgesellschaft, Berlin.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus gracilis
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: gracilis Gosse, 1864
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: taxon inquirendum; keine Angaben über Schuppen und Stacheln, daher nicht wieder erkennbar
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Gosse,P.H. 1864. The natural history of the hairy-backed animalcules (Chaetonotidae). The Intellect. Observer 5: 387-406.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus hermaphroditus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: hermaphroditus Remane, 1936
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: taxon inquirenda; jegliche Abbildung und Beschreibung fehlt!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Remane,A. 1936. Gastrotricha, In: H.G. Bronns, ed. Klassen Ordnungen des Tierreichs, Band 4, Abteilung II, Buch I, Teil 2, Lieferungen 1-2. Akademie Verlagsgesellschaft, Berlin.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus lancearis
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: lancearis Tretyakova, 1992
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Dorsal scales: dorsolateral 3 Bündel mit leicht gebogenen, abspreizbaren Stacheln mit je zwei Nebenspitzen; (Hals: 3 x 130-140µm, 2 x 35-38µm; Mitte: 3 x 80-90µm; Hinten 2 x 60-70µm)
Dorsal scales: Dorsolatereal 9 Paar s-förmig geschweifte Stacheln mit 2 Nebenspritzen in 3 Bündeln (Hals: 3 x - erste zwei 180-182µm, dritte 40µm; Mitte: 4 x 130-132µm; Hinten: 2 x 60-72µm)
Dorsal scales: Hinten: 2 stumpfe Lappen mit kurzem Stachel (17-20 µm) mit Doppel- und Nebenspitze; auf einer Seite 12, auf anderer Seite nur 3 z.T. riesige Stacheln mit Doppel- und Nebenspitze; 3 dünne Stacheln am Hinterende (50µm)
Ventral scales: 4-5 Paar Zilienbüschel, sonst nackt
Oecology: über Faulschlamm, Lemnatümpel
Similar species: durch asymmetrische Stacheln einzigartig
Dorsal scales: Auf den Hinterlappen ein Paar langer, auf Schuppenrudimenten sitzende Stacheln (63-68 µm; 1/4 Körperlänge); 2x7 Stachelbündel mit proximaler Nebenspitze; Kopf 2x mit 1 bzw 3 Stacheln; Rumpf 5x mit 4-7 Stacheln; Stachelanzahl und -länge (60.76µm) variiert
Ventral scales: 5 Paar Wimpernbüschel
Oecology: semiplanktisch über Schlamm
Similar species: evtl. synonym mit
D. (D. ) ornatus
; Unterschied Stacheln
Particularities: sehr viele gleichmäßiger verteilte Stacheln
Dorsal scales: Kopf zwei Ringe mit je 6 Stacheln mit einer Nebenspitze (10-15 µm); Körper 4 Paare Stachelgruppen mit je 5 Stacheln mit zwei Nebenspitzen (69-115 µm); Hinten zwei Paare Stacheln mit zwei Nebenspitzen (18µm); zwei Paar einfache Stacheln
Dorsal scales: neun Paare Stachel-Gruppen; Stachelzahl von vorne: 1 - 1 - 2 - 5 - 5 - 3 - 1 - 1 (58-75 µm); erste beiden Gruppen einfach, sonst zwei Nebenspitzen; auf Schuppenrudimenten; Schlägerartige unpaarige Struktur am Hinterende (59x17 µm), kann fehlen (vielleicht epizooisch)
Oecology: zwischen Wurzeln; Brasilien
Similar species: D. (D. ) ornatus
hat weniger Stachelgruppen und keinen “Schläger”
Dorsal scales: terminal 2 dick Stacheln (78-85 µm), distal mit Doppelspitze, an der proximalen Nebenspituze stark abgewinkelt; 8 Paare Stachelbündel: Kopf 1x; Hals 2x; 6 Paar Rumpf 3-5 x; längste stacheln 84-86 µm; am Kopf 3 weiter Stacheln, mittlere ohne Nebenspitze
Dorsal scales: No scales; 5 pairs of non-crossing elastic spines projecting above the body; two spines each starting ventrally
Ventral scales: no scales; 3 pairs of long jump spines (100 - 240 mm); posterior ones overhang the body by one body length and cross each other
Oecology: over mud, formerly frequent, meanwhile rare
Similar species: H. festinans
: lower number of jump spines
Particularities: Setae stand on clear papillae
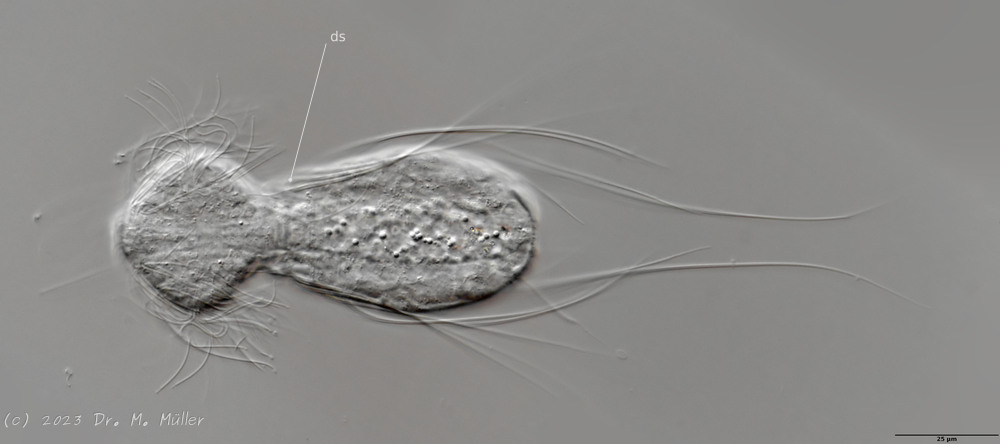
Haltidytes crassus was a relatively common species at the time of its first description. Since then - probably due to the influence of industrial agriculture - this pretty gastrotrich has become very rare. The main characteristic of H. crassus are the five dorsal and three ventral “jumping spines”, with which the animal catapults itself out of the danger zone when threatened.
Dorsal; ds: dorsal spines (5 pairs).
The long, caudal and hair-shaped last ventral pair of spines, which far overhang the body and cross each other in resting position, is conspicuous.
Three ciliary rings on the head are used for swimming locomotion, while the ventral ciliary bands typical of gastrotrichs are used for gliding over the substrate.
Ventral; cr: 3 ciliary rings for locomotion; js: jump spines; cs: long caudal spines; se: seta on papillae.
The posterior palps - as in all members of the genus Hatidytes - are on distinct papillae.
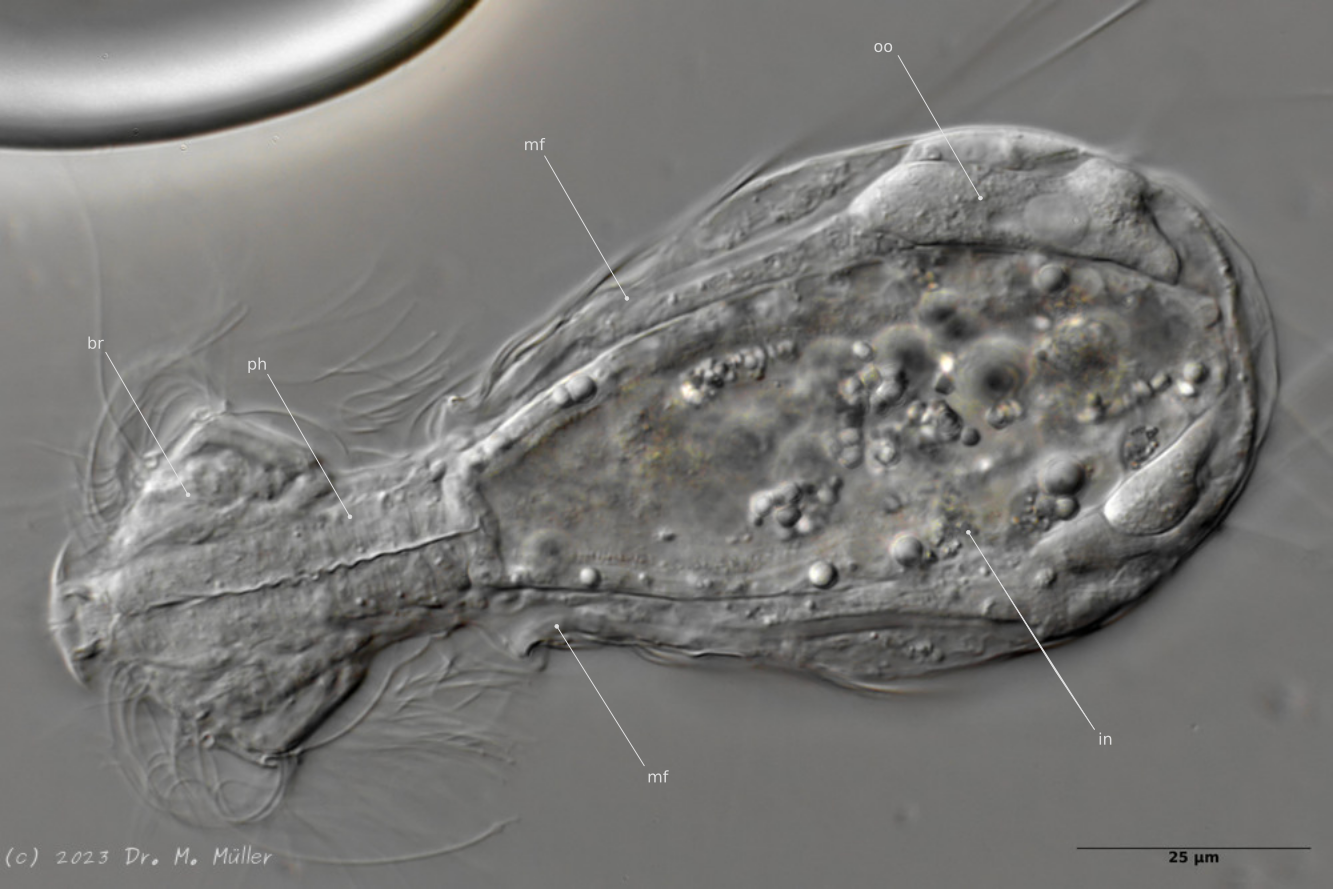
Median; br: “brain”; ph: pharynx; oo: mature ovum; in: “intestine”; mf: muscle fibers that move the jumping spines.
The internal anatomy has some peculiarities. For example, distinct muscle fibers can be seen that couple to the attachment sites of the jumping spines. Adhesive glands and the associated adhesive tubes are not present in this genus.
The typical movement and behavior of the animals can be seen in the following YouTube video:
Dorsal scales: no scales; dorsolaterally 3 groups each of 3 + 2 + 2 jump spines, which protrude even in resting position and slightly overhang the body
Ventral scales: no scales; a pair of long jumping spines (140-144 µm), surpassing the body by 3/4 of its length and crossing each other
Oecology: About digested sludge
Similar species: H. crassus
: more ventral spines, less lateral spines
Heterolepidoderma macrops is one of the most common and widespread Heterolepidoderma species and was certainly often confused in the past with
H. ocellatum
from which it differs - apart from some details that are difficult to see - mainly by the larger number of scales:
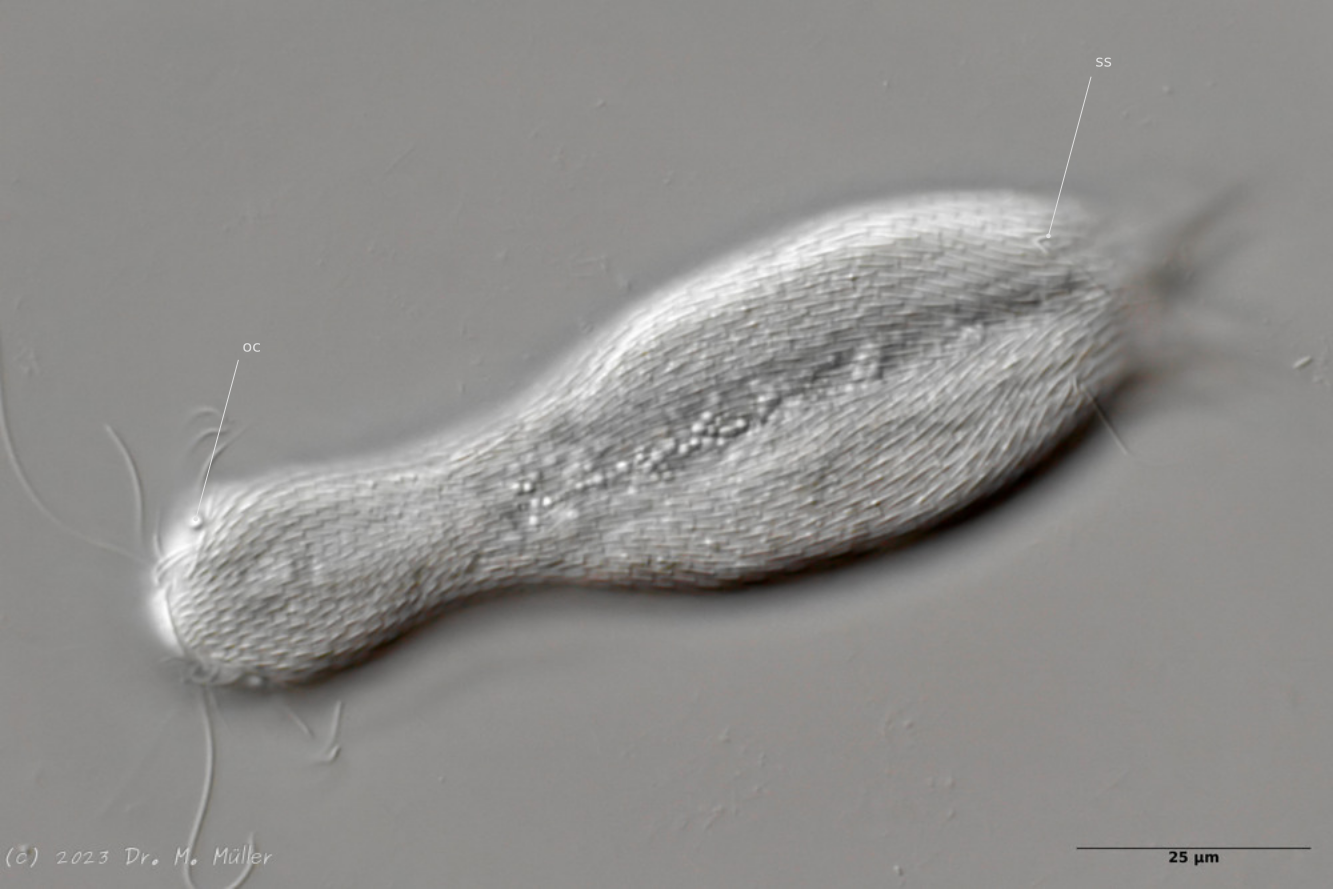
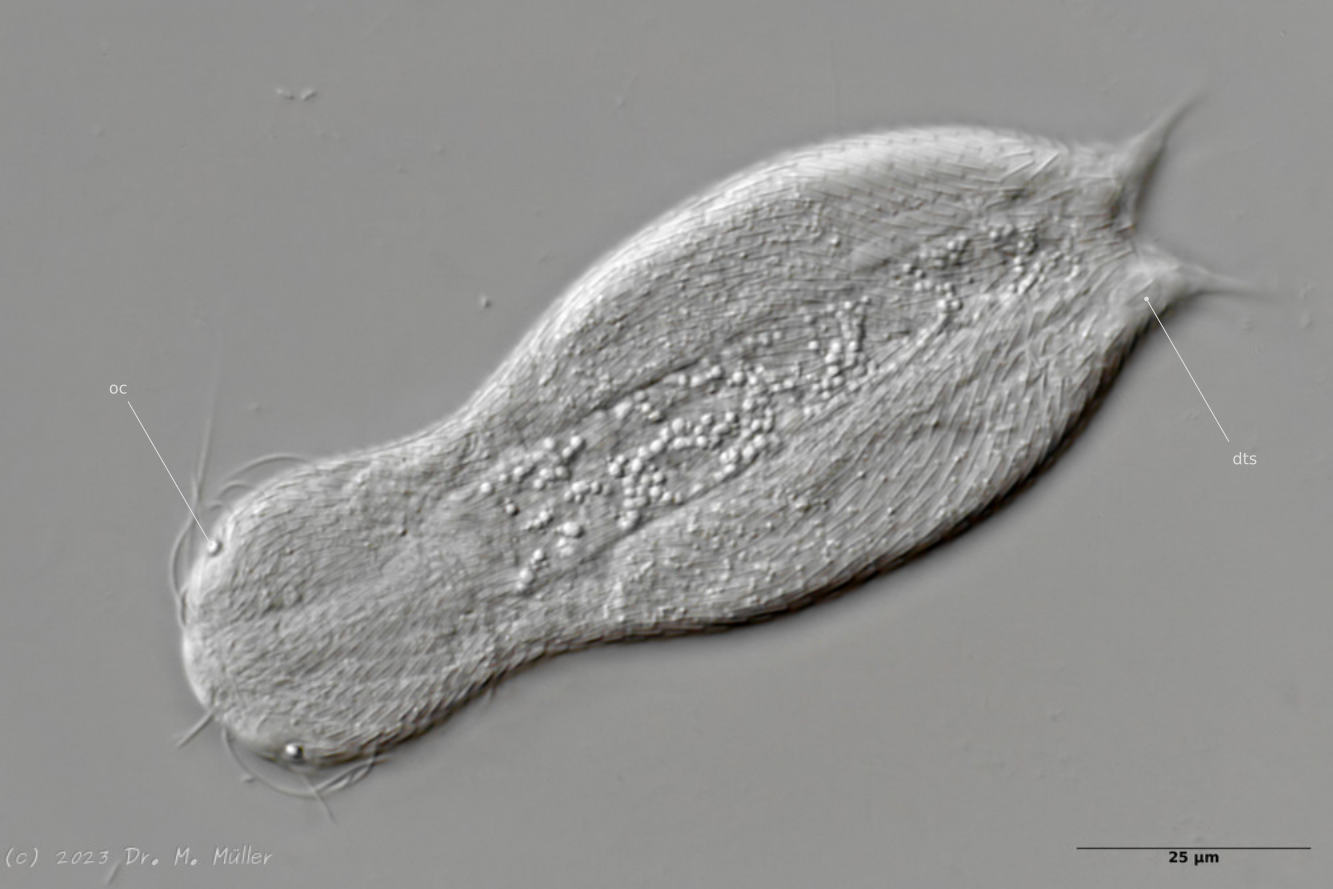
Dorsal view; oc: pseudocells; ss: sensory scales.
In most animals, the pseudocells resembling eyes are very conspicuous, but they have no sensory function. The posterior sensory hairs are located on small, triangular “sensory scales”.
Dorsal with focus on the dorsal terminal scales (dts).
The toes of the animals are unscaled. The scale ends with a pair of relatively large keel scales at the base of the toes.
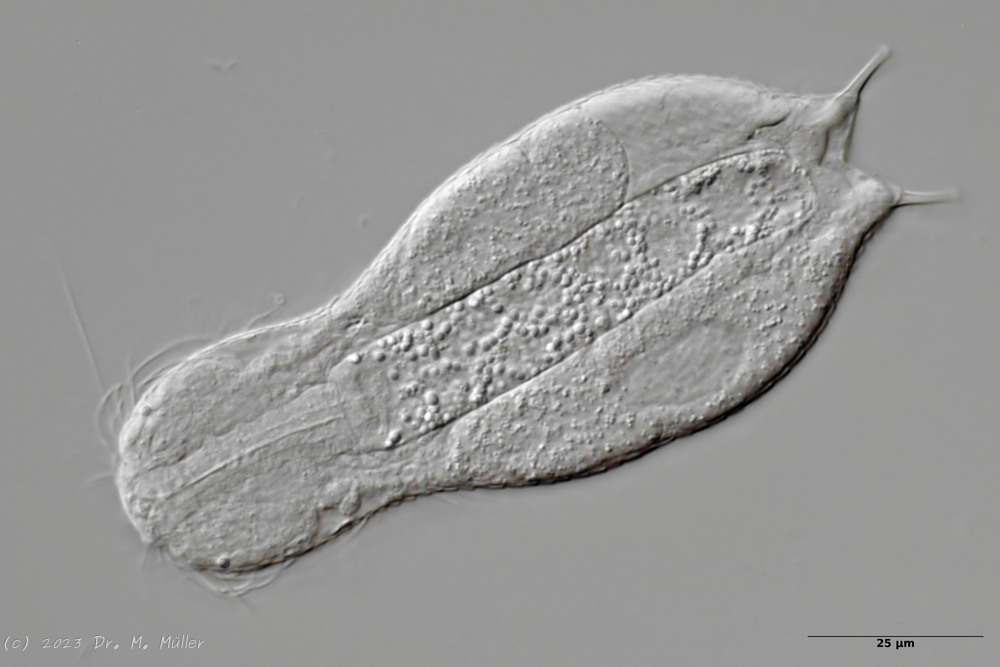
*Cross section
In the section you can see that the small scales - in contrast to
H. ocellatum
- do not bear a distal process. The pharynx is slightly swollen on both sides.
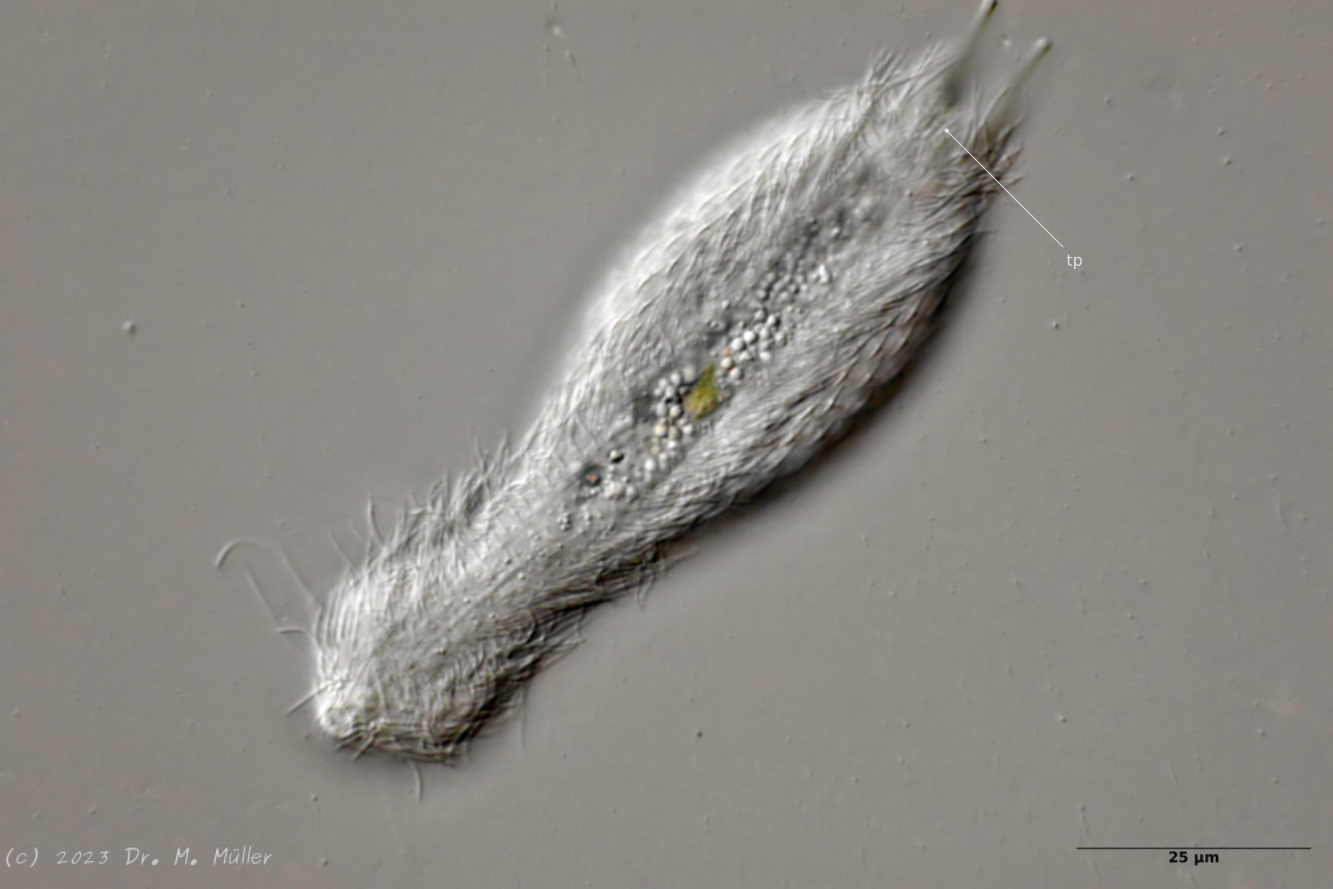
Central view with terminal plates (tp)
Ventrally, the animal bears only a pair of small, oval terminal plates. The remainder of the ventral interfield is unscaled.
The adhesive tubes are thickened terminally.
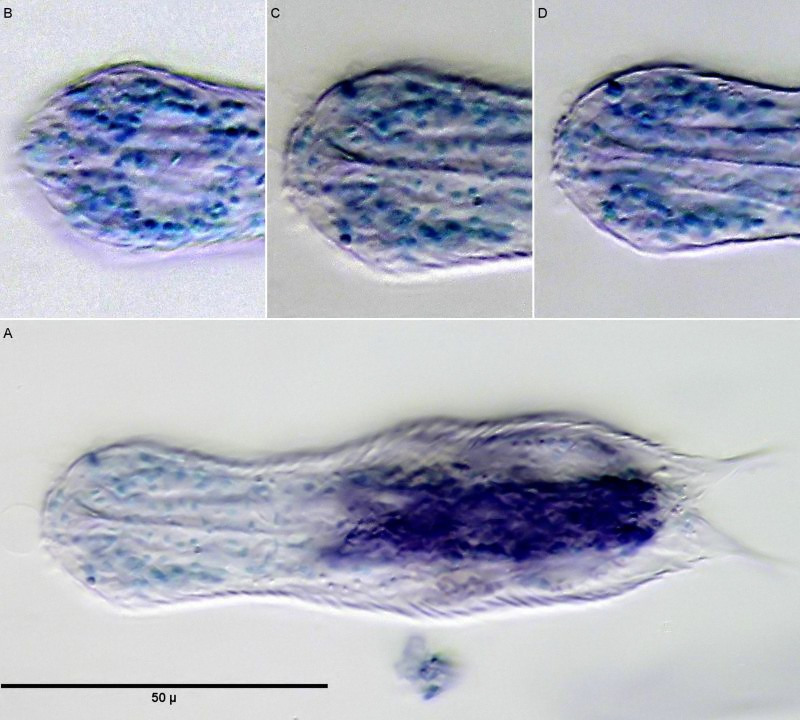
Staining of nuclei with methyl green; B C D: Nuclei of the brain; A: overview.
If H. macrops is stained with a selective nuclear dye like methyl green, only the nuclei of the animals are stained. This gives a good overview of the structure of the brain. Clearly visible in subfigure B is the nucleus-free area dorsal to the pharynx, which is referred to by (Zelinka, 1889)and (Remane, 1936)as the “dorsal commissure”, i.e. a nucleus-free connection of the two cerebral hemispheres made of nerve fibers, and which is also confirmed in more recent studies (Schmidt-Rhaesa and Holger Rothe, 2014). Lowering the focus to the midplane of the pharynx (subimage C), one recognizes the pharynx consisting of a few cells surrounded on both sides by brain cells. Ventrally, no connection of the two cerebral hemispheres can be seen below the pharynx. Thus, the brain does not completely enclose the pharynx and delimits the animals to the Cycloneuralia. The already unstained conspicuous pseudocells prove to be stainable with methyl green - a strong indication that these structures are the prominent nuclei of the sensory neurons of the tactile hairs.
Scale preparation with eosin.
The scale shape of the small dorsal keel scales of H. macrops cannot be judged exactly in the scale compound on the living animal. A more precise [scale preparation](/technique/scale preparation) clearly shows the oval shape of the scales, which - in contrast to
H. ocellatum
- do not wear a distal tip.
After a phase with parthenogenetic reproduction, the animals enter the so-called “postpathenogenetic phase”. In this stage of life, the animals reform into hermaphrodites and simultaneously develop eggs and sperm produced in bundles ventrally to the side of the intestine - they reform into true hermaphrodites.
In H. macrops the spermatoids are spindle-shaped. At the same time, a bipartite, so-called X-organ with unknown function is formed. Here the X-organ consists of two spherical, secretion-filled cysts connected by a bar (or connecting duct).
Heterolepidoderma majus is a very widespread, slender and elegant belly-hardy.
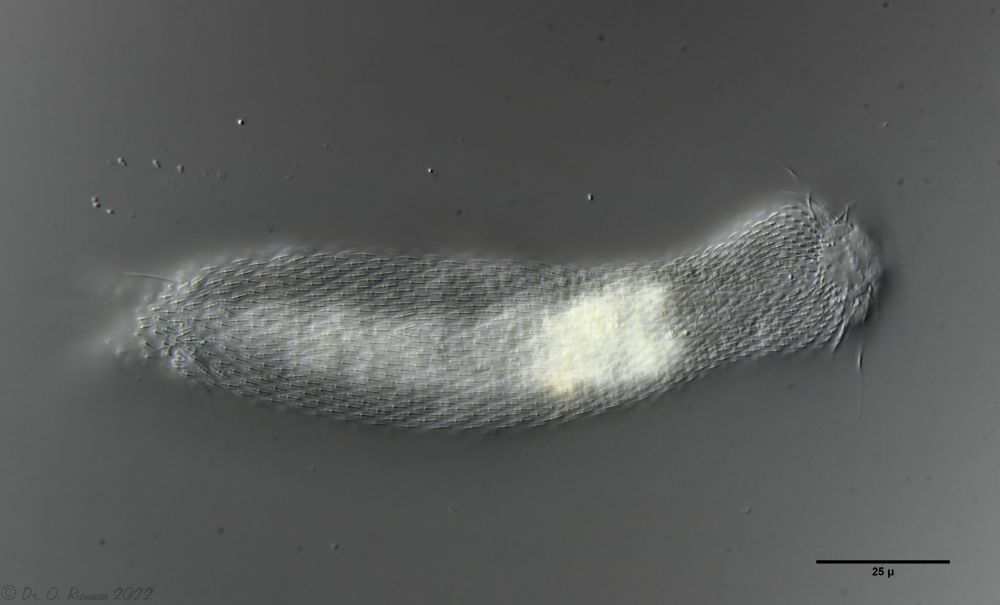
H. majus: dorsal scaling ( Ole Riemann)
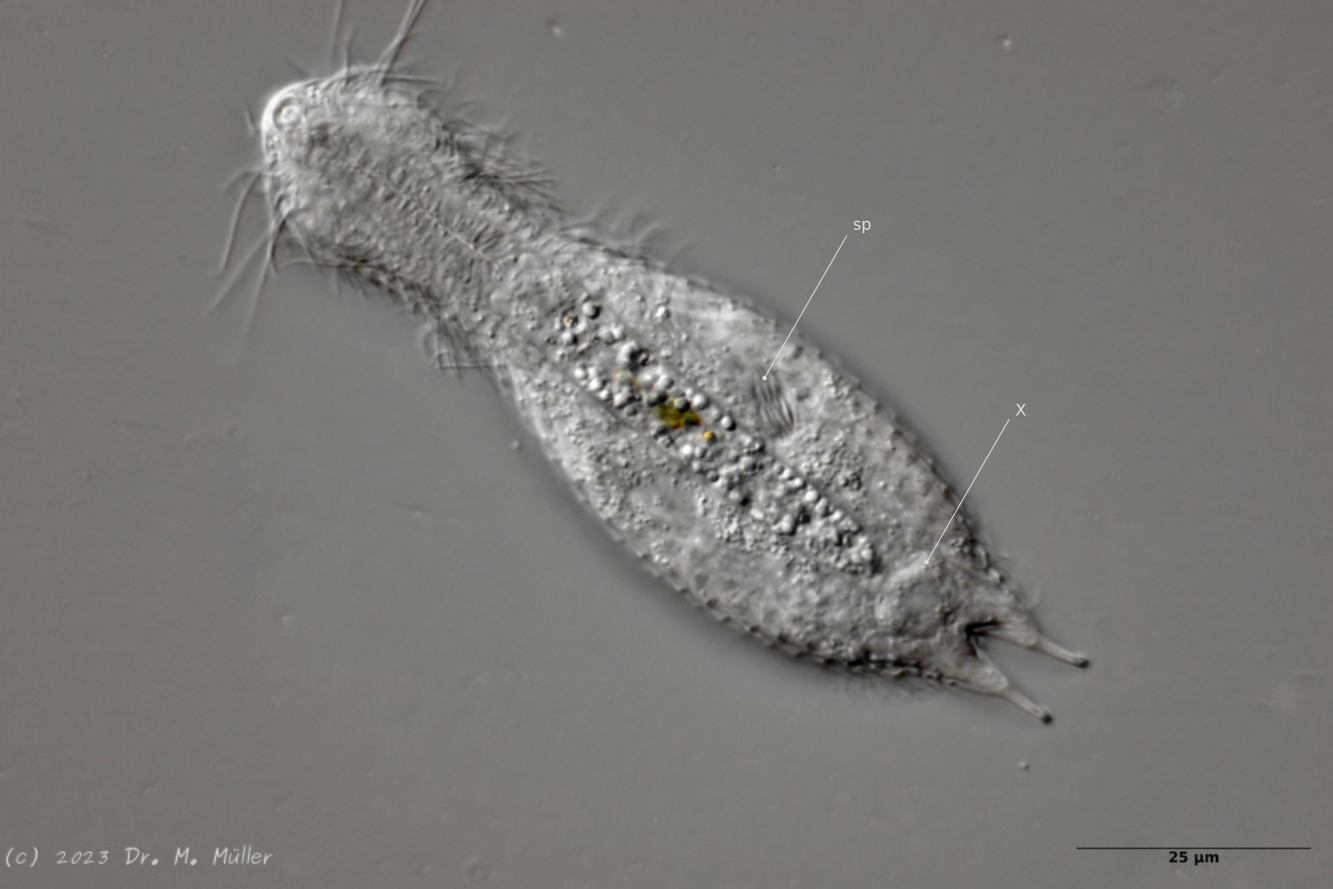
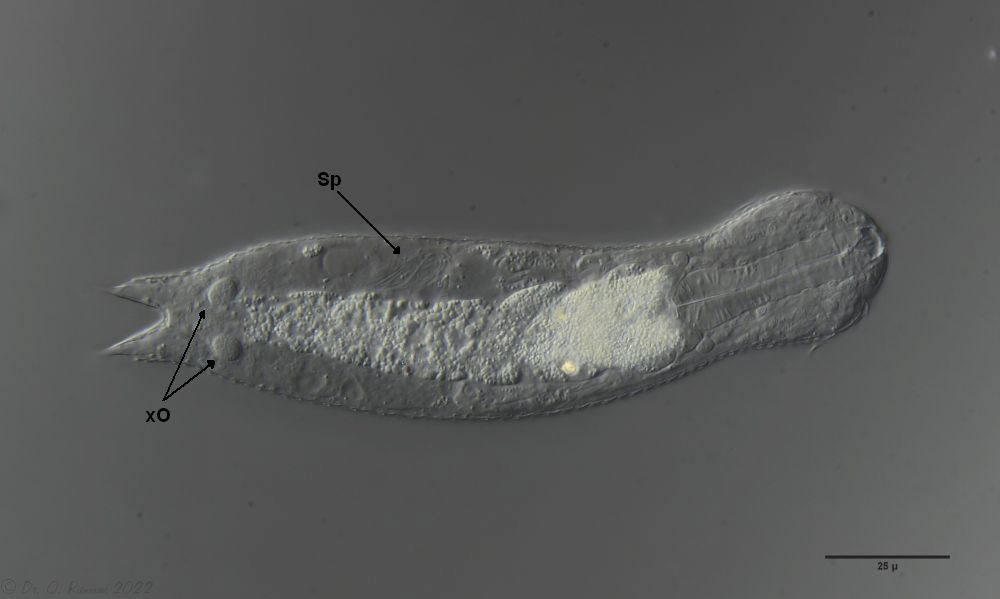
One of the animals observed was in its post-parthenogenetic (hermaphroditic) phase and showed a sperm bundle in addition to the conspicuous bipartite X-organ at the posterior end:
H. majus: x-organ (xO) and sperm bundle (Sp)( Ole Riemann)
The scaling at the base of the toes is typical for the species. The sensory hairs arise from special scales:
H. majus: Scaling of the base of the toes ( Ole Riemann)
The ventral intermediate field is occupied by small, elongate keel scales and bears two terminal keel plates:
H. majus: Scaling of the ventral interfield
Another feature that makes the species distinctive is the demarcated “stomach ring” on the foregut:
Similar species: Ichthydium monolobum was included in this species
Particularities: Very characteristic furca
The species I. (F.) forficula was united some time ago with the somewhat smaller species I. monolobum into a single species. A main characteristic of this species are the long, curved and thin glue tubes, which give the animals a “tweezer-like” appearance:
I. forficula: dorsal with pincer-like adhesive tubes
On the ventral side the animal carries two distinct terminal plates.
I. forficula: ventrale Terminalplatten
The lash bands are broken up into individual clumps of extremely long lashes.
I. forficula: Ciliate tuft
In the side view, you can see the enormous length of the locomotive eyelashes:
Dorsal scales: only triangular basal scales of terminal setolae
Ventral scales: naked except for two rectangular, unkeeled terminal plates; rows of cilia on trunk resolved into 4 pairs of ciliate tufts; bands of cilia united on head
Oecology: In sphagnum / bog
Similar species: I. (F. ) skandicum
: Additional dorsal scales at posterior end
Particularities: Very long dorsally visible cilia (27µm); very rare, only a few specimens in Italy and Sweden so far.
Ichthydium (Furficulichthys) tanytrichum is a very rare belly-harper, which since its first description in Italy in 1982 (Balsamo, 1982)has only been reported from Sweden - as a single specimen (Kånneby et al., 2009). The here shown finding from the Austrian Lauchseemoor is the third worldwide known occurrence of this species.
With a size of about 130µm the animals of the found population are about the same size as the animals found in Sweden. The Italian animals of the first description were significantly smaller (ca. 74µm). Already Kånneby expressed the suspicion that the first description was based on juvenile animals, because the adult animal found by him was clearly larger with 127µm. This would also explain the somewhat different proportions to the first description.
Median cut
The lack of a continuous scaling and the flexible cuticle are characteristics of the genus Ichthydium. The typical pincer-shaped furca of the animals places I. tanytrichum clearly in the subgenus Furficulichthys, within which the species can be clearly defined by body shape and lack of scaling.
Dorsal: “sensory scales” from which the tactile hairs arise.
Other species characteristics are the triangular “sensory scales” from which the posterior tactile hairs arise. The anterior pair of tactile hairs sits on small papillae. Further scales are not present on the dorsal side.
Ventral: In the ventral region, the cilium bands are resolved into single tufts.
On the ventral side the long cilia are noticeable, with which the animal moves. With a length of about 25µm they give this species its name (“tanytrichum” = “long haired”). Typical for this species is, that these locomotive cilia are not organized in two continuous cilium bands - as otherwise common in gastrotrichs - but stand in single cilium tufts. Terminal are faintly recognizable the two large rectangular terminal plates.
One of the animals found had reached its postpathenogenetic (hermaphrodite) phase and was already carrying an X organ and sperm bundle:
L. minor chaetifer: above dorsally with smooth scales typical of Lepidodermella;
middle in cross section with X-organ and sperm bundle;
below ventral scaling with distinct terminal plates ( Ole Riemann).
In Sima Marsh the somewhat divergent subspecies Lepidodermella minor chaetifer was found, differing from the species L. minor minor by a pair of spines at the base of the toes:
L. minor chaetifer: the pair of spines at the base of the toes is characteristic of the subspecies chaetifer
Another striking feature of this species is the single tooth projecting asymmetrically from the oral cavity:
Dorsal scales: 7-9 rows of 25-30 very differently shaped, spikeless, strongly overlapping scales; head and trunk scales elongated oval with triangular / broadened front end (12 x 10 µm); neck scales transversely oval, short (5 x 9 µm); hypostomium: two cusps
Ventral scales: ventral intermediate field with more than 20 transverse claspers below pharynx and 3-5 rows of quadrangular rounded, overlapping scales; 2 large terminal plates and some small, round scales; ciliary bands anteriorly mostly united
Oecology: Frequent, found everywhere
Particularities: well delimited by rectangular scale plates and large terminal plates; very variable and well studied species
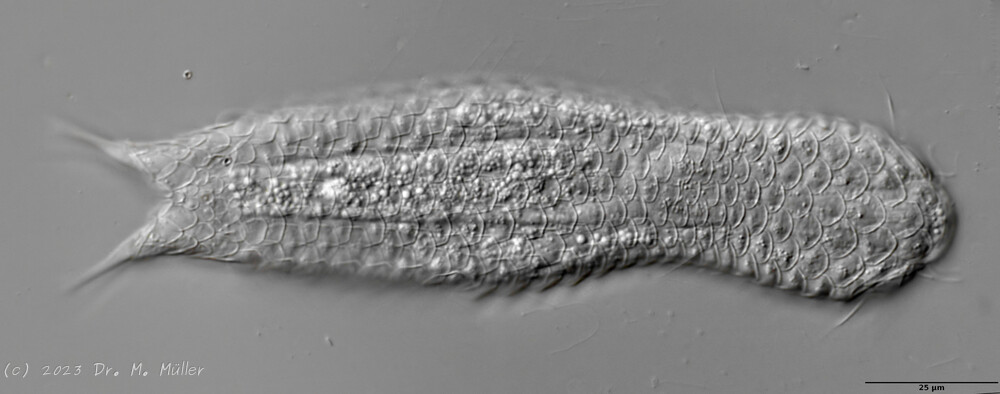
Lepidodermella squamata is one of the best-studied gastrotrichs, as it is kept in culture in many laboratories around the world. However, the “cultivated forms” often differ significantly from the “wild forms”, as the animals, with their enormous generation rates of around 50 per year, can react quickly to the adaptive pressure of the habitat. L. squamata is therefore considered a very variable species. The photos shown here are of wild forms.
Dorsal scales
Ventral scales; large terminal plates; rectangular braces under neck
L. squamata is one of the many species of gastrotrichs in which the glandular ducts on the surface of the pharynx are clearly visible. These ducts connect individual gland cells in the pharyngeal wall and channel the secretions of the glands into a common excretory duct that pours into the animal’s oral cavity:
Width of the head ( five lobes, broad ): 38 µm - 40 µm
µm
Length of the furca:
Adhessive tubes:
Pharyx ( cylindrical ): 35 µm
Diameter of the mouth ( around ): unknown
Dorsal scales: posterior setolae on small keel scales; terminal spine 45µm; spines in 5 groups of one short ventral spine (10-15 µm) and one longer lateral spine (ca. 40 µm); anterior and last group unpaired, i.e. 8 spines per side
Ventral scales: naked; only one pair of eyelashes; eyelash tufts on head typical
Oecology: plant-rich pools; over mud; rare
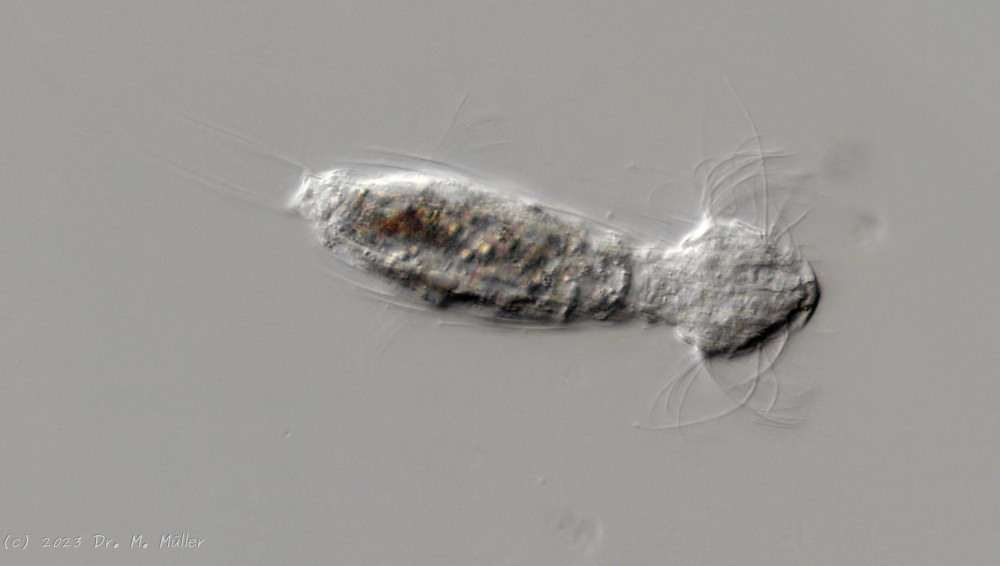
Setopus bisetosus (Thomson, 1891) - a formerly common species - has become very rare due to the progress of industrial agriculture and the accompanying drainage. Therefore, it always pleases me to find a specimen of this species.
free swimming copy
Unfortunately, I could not completely still the animal, so the photo quality was not optimal.
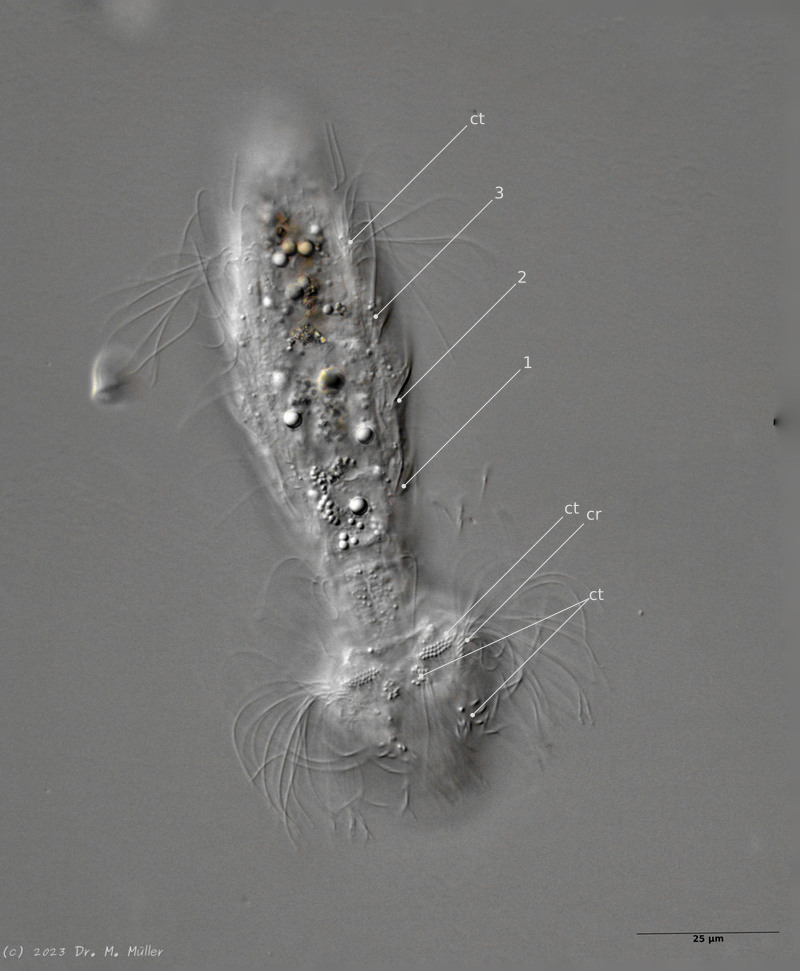
The semi-planktic Setopus species usually do not have continuous ciliary bands and move swimming with an (almost completely closed) ciliary ring on the head. In addition, some ciliary tufts are present ventrally on the head. On the body itself, except for a few isolated cilia, the ciliary bands are reduced to a pair of ciliary tufts on the abdomen.
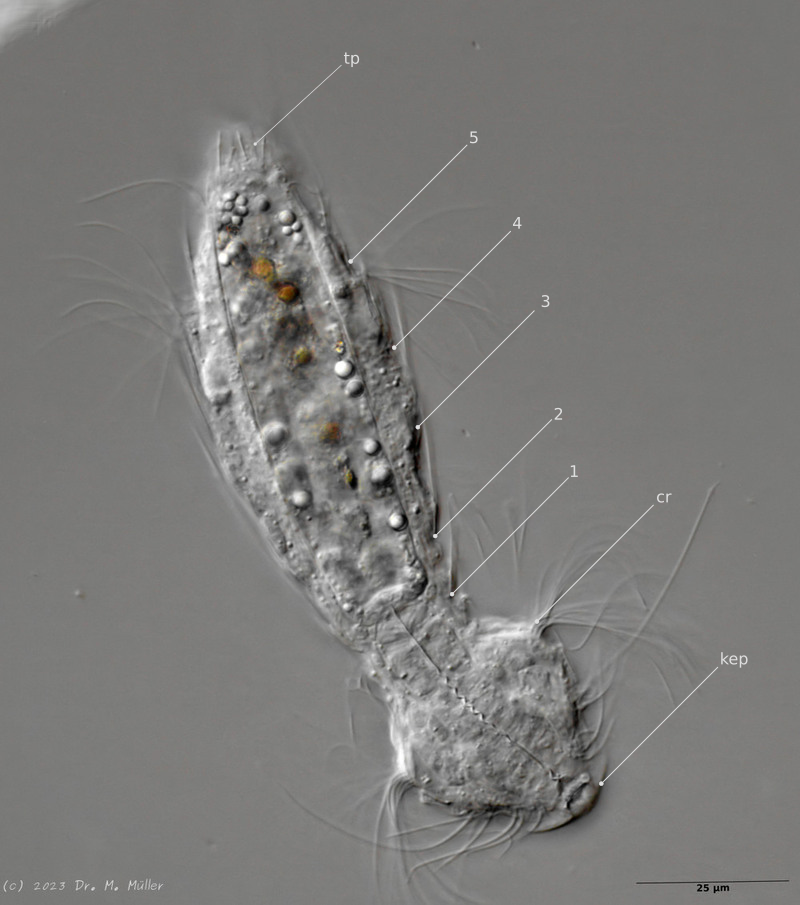
In median section, two pairs of keeled terminal plates can be seen at the posterior end of the animal. These terminal plates are not mentioned in the literature.
Laterally five pairs of spines are visible.
Median section with terminal plates (tp) and head shield (kep); 1..5: lateral spines; cilia ring (cr).
Typical is the broad and strong head shield (kephalion) of the animals.
Dorsal scales: Terminal spines with secondary apices on triangular scale rudiments; unequal in length (27-30 µm; 43-55 µm); numerous dorsolateral spines with secondary apices on scale rudiments; 21 per side.
Oecology: rare; bog lake
Similar species: S. chatticus
: no scales and fewer shorter spines
Particularities: scaled ventral interciliate field
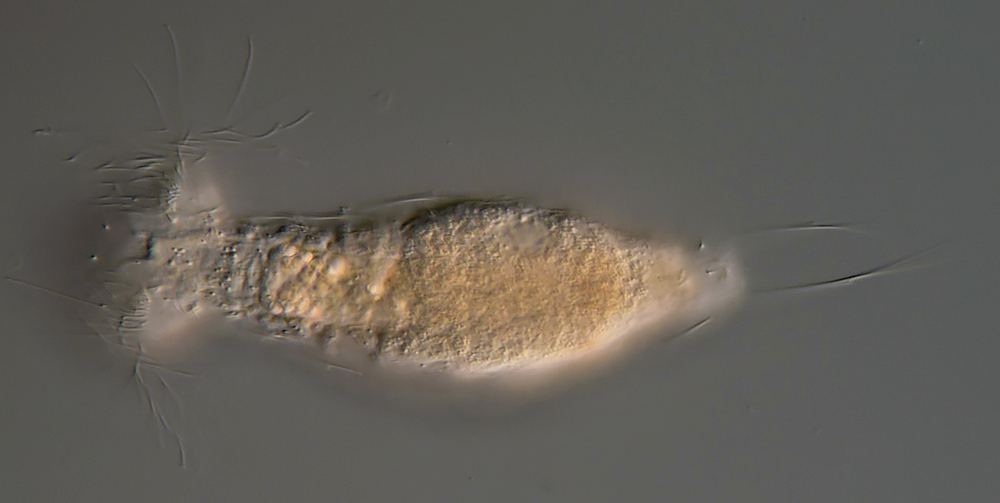
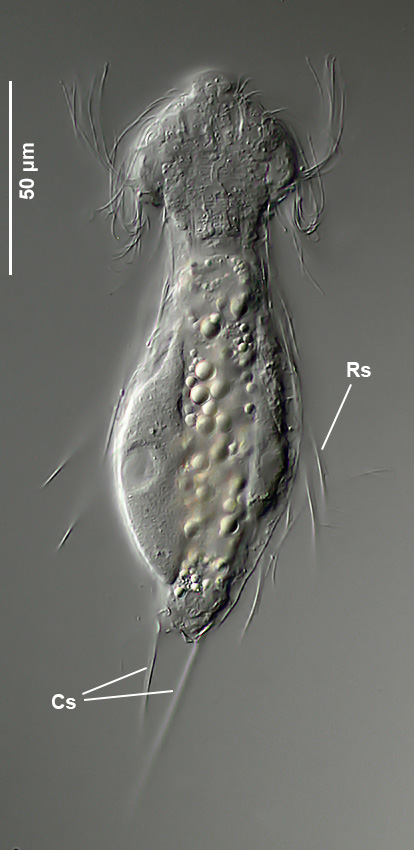
Setopus tongiorgii has been found so far only in Italy and some locations in Germany and is - like all Setopus species - relatively rare.
Overview with focus on the ring of eyelashes on the head ( Dr. Stephan Krall).
Transverse section; Rs: trunk spines; Cs: caudal spines ( Dr. Ole Riemann).
In the cross section the lateral spines with their secondary spike are well visible. Terminal are two caudal spines of unequal length, which also carry a secondary tip.
Scaling of the ventral intermediate field ( Dr. Ole Riemann).
Ventrally, the small, roundish scales and the keeled terminal plates of the ventral interciliar field can be seen. So far only in S. tongiorgii scales were found in this area. This feature is therefore of special importance for the diagnosis of the species.

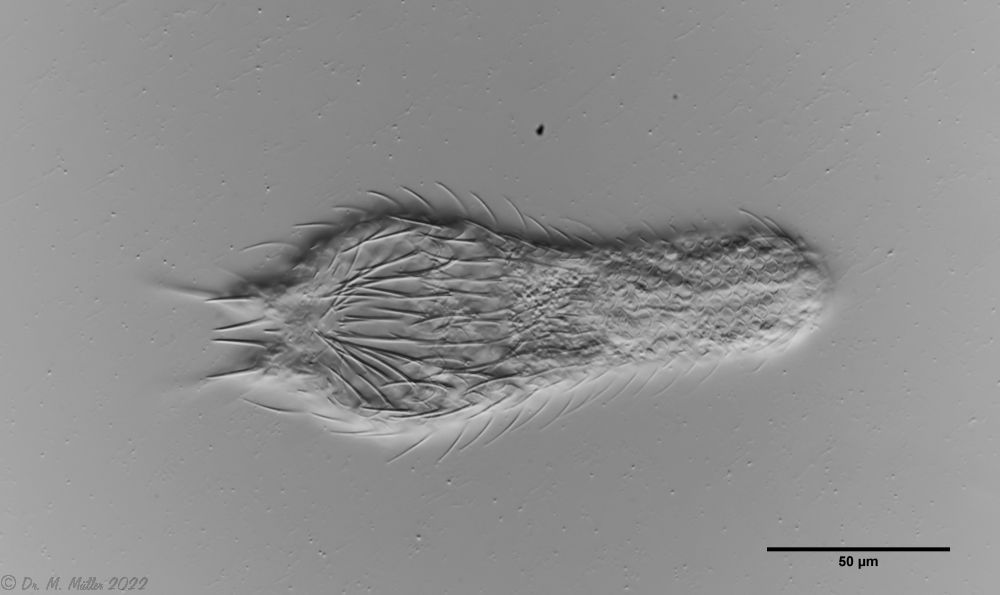
 typical scales of an Aspidiophorus, here A. squamulosus.
typical scales of an Aspidiophorus, here A. squamulosus.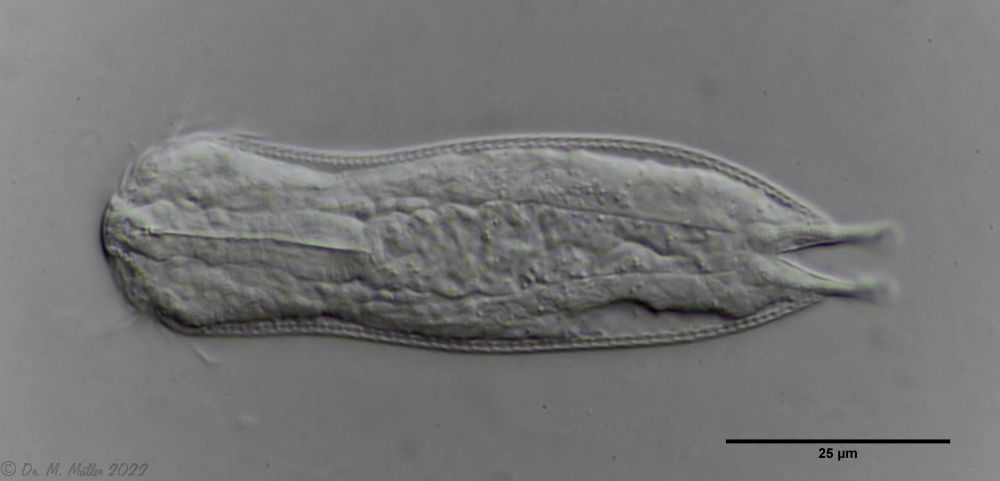




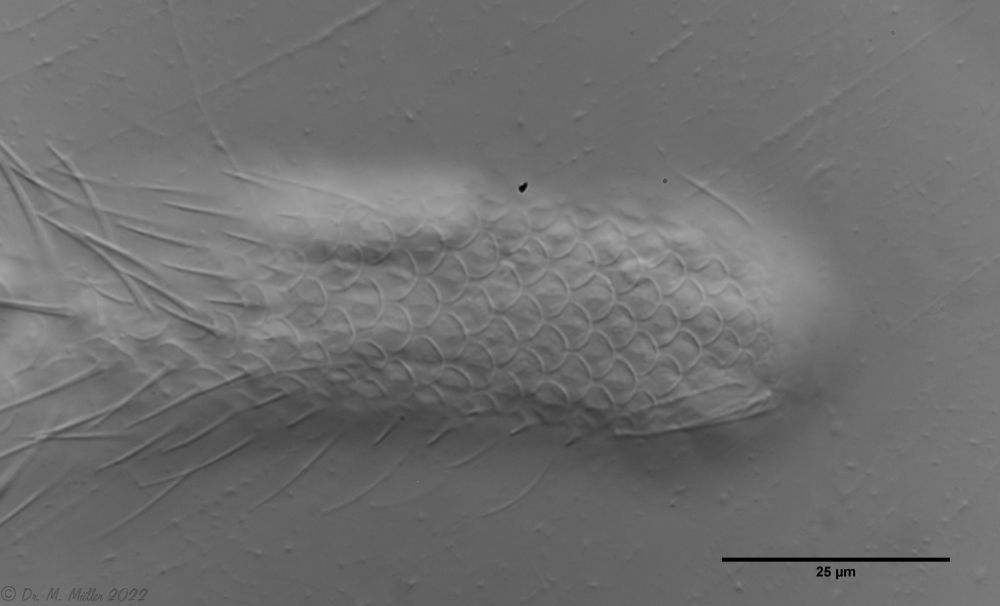
 Backscales
Backscales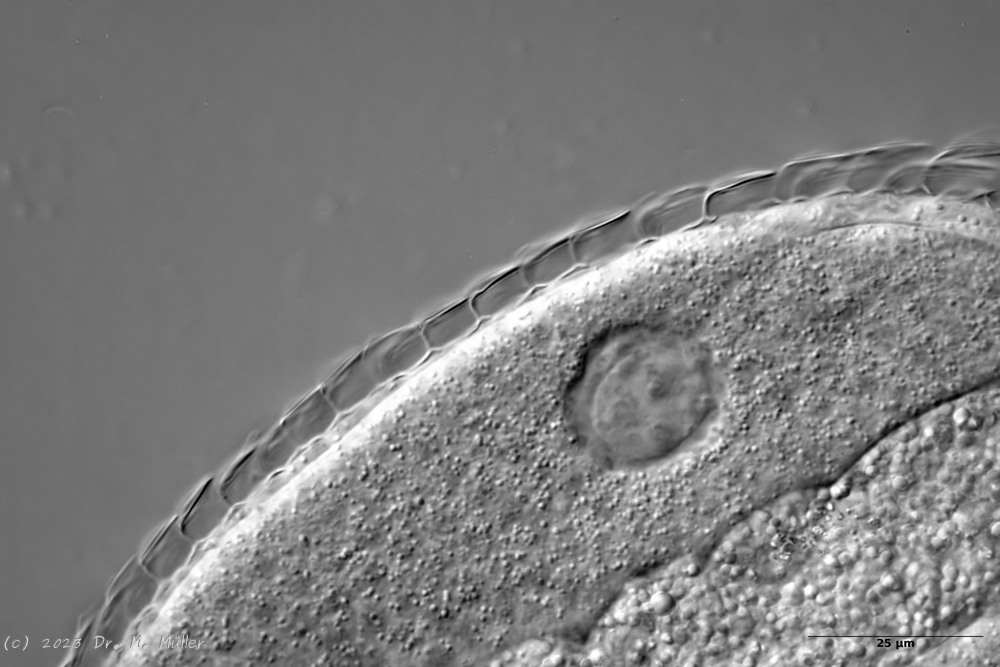
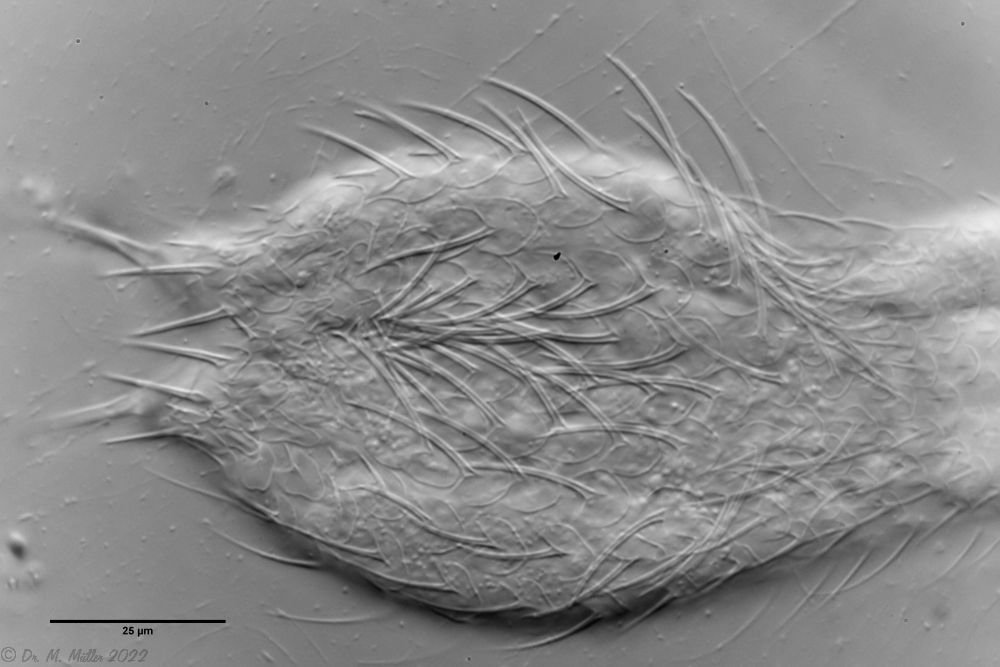
 Cross section scale stems
Cross section scale stems







 Ch multispinosus: ventral
Ch multispinosus: ventral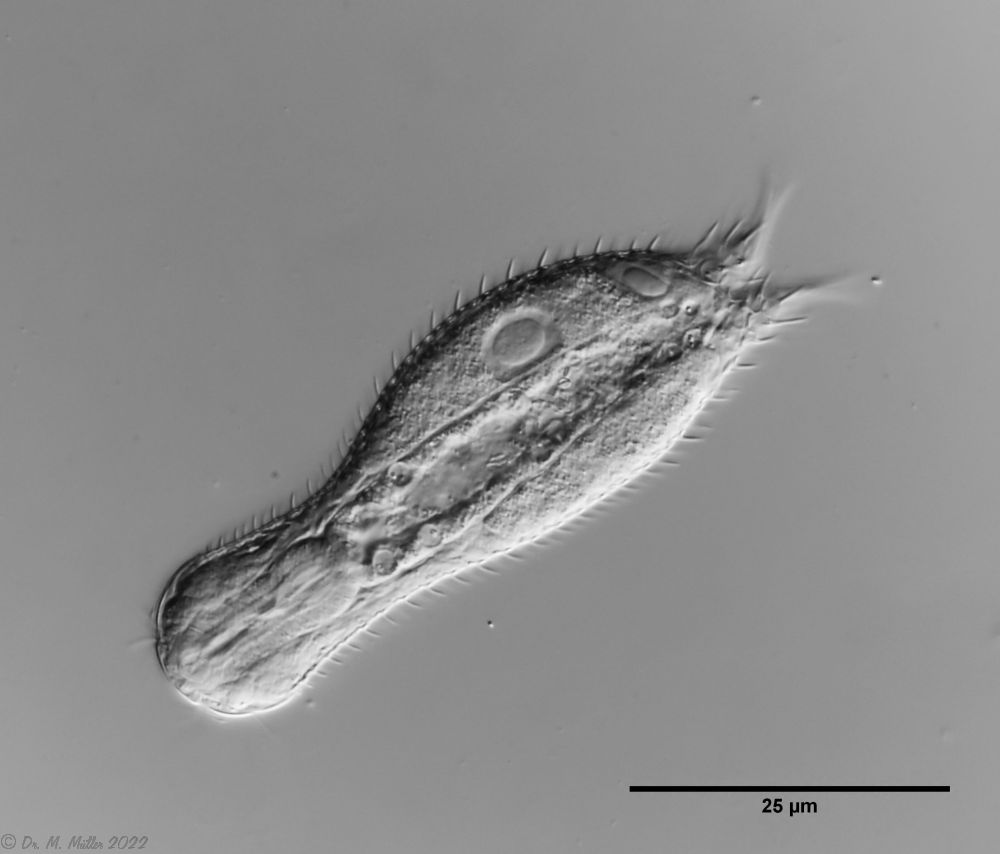 Ch multispinosus: cross section
Ch multispinosus: cross section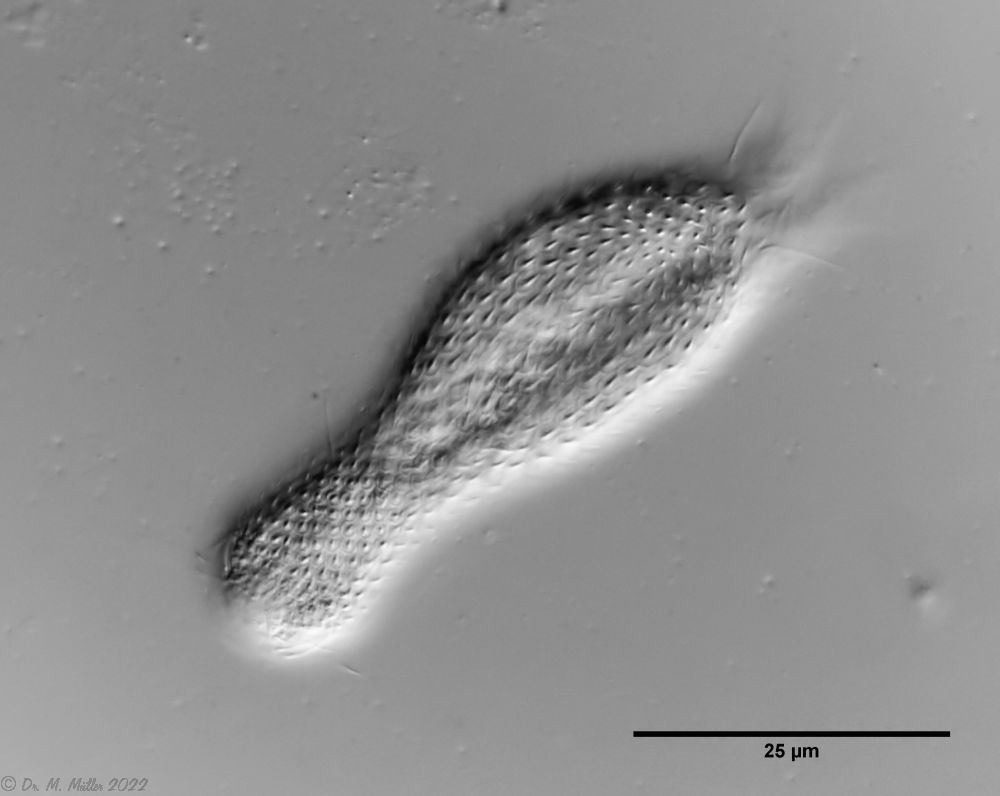 Ch multispinosus: dorsal
Ch multispinosus: dorsal *Typical Chaetonotus (Hystricochaetonotus) - here
*Typical Chaetonotus (Hystricochaetonotus) - here
 Ch. murrayi: cross section
Ch. murrayi: cross section Ch. murrayi: dorsal
Ch. murrayi: dorsal Ch. murrayi: focus on the spine tips
Ch. murrayi: focus on the spine tips Ch. murrayi: dorsal scaling of the head / neck area
Ch. murrayi: dorsal scaling of the head / neck area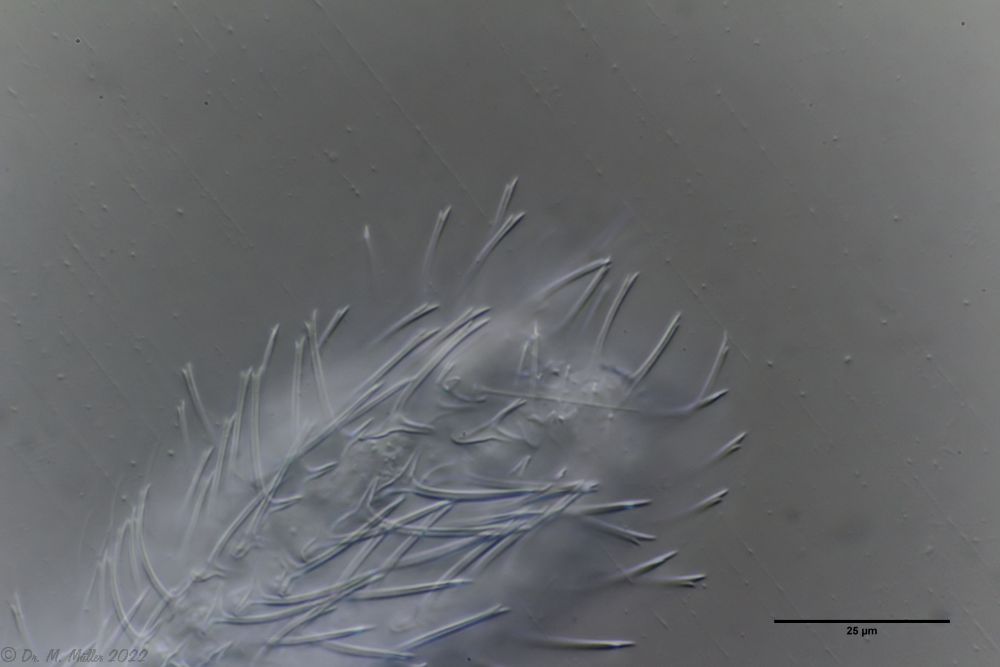 Ch. murrayi: dorsal scaling of the hind end
Ch. murrayi: dorsal scaling of the hind end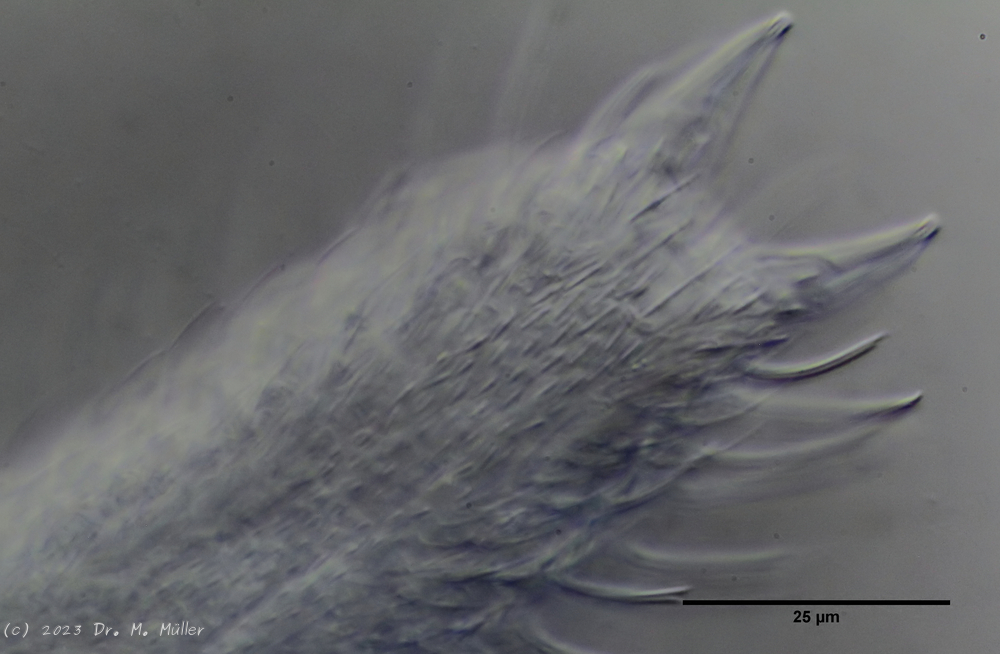 Ch. murray: Scales of the ventral intermediate field
Ch. murray: Scales of the ventral intermediate field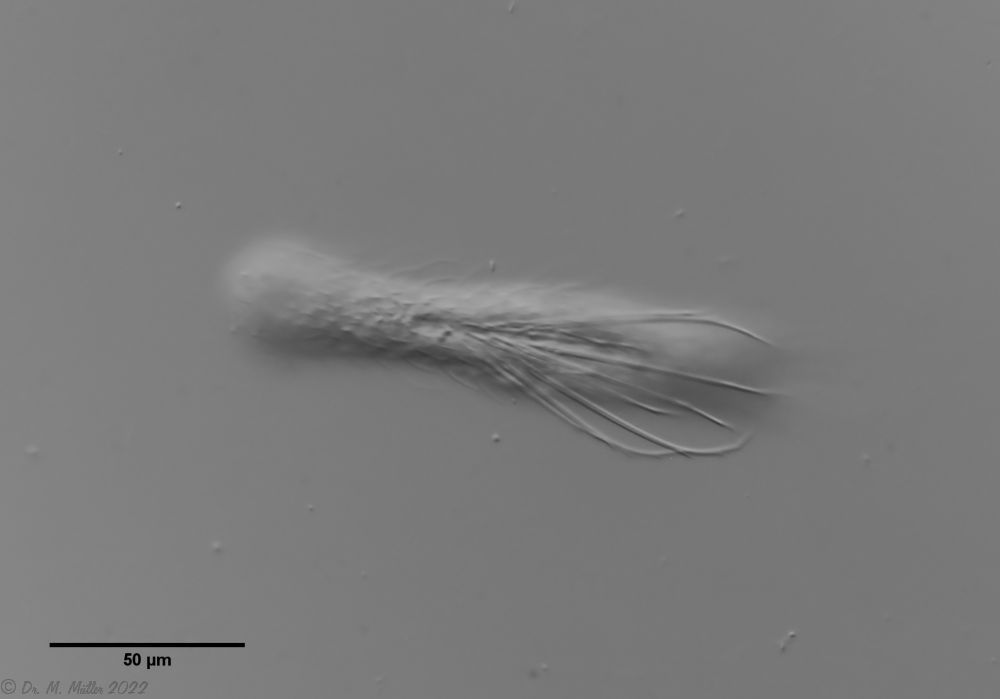 Ch. novenarius: dorsal
Ch. novenarius: dorsal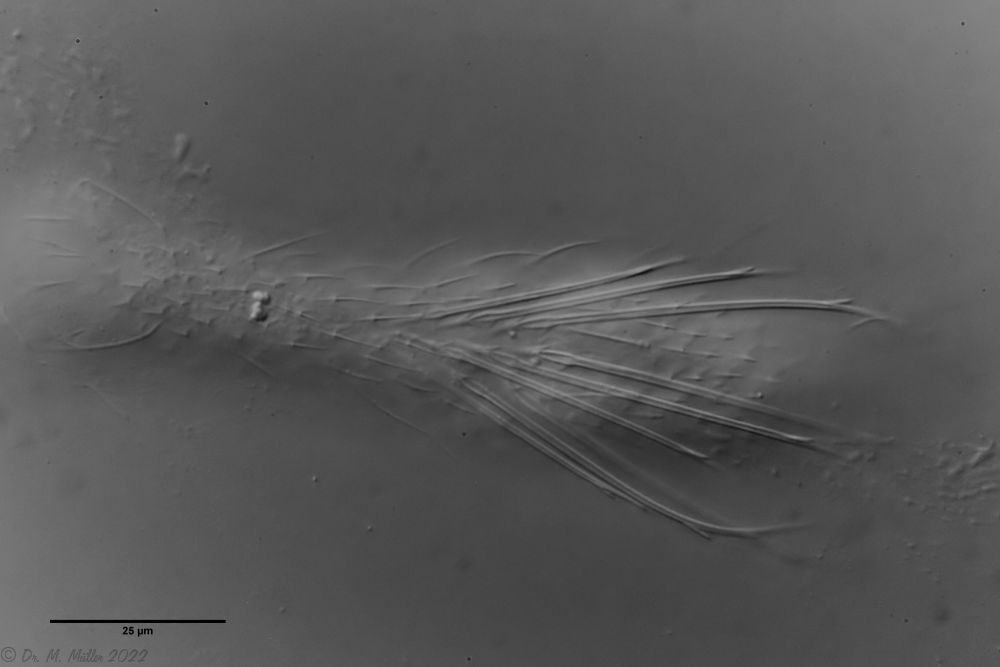 Ch. novenarius: dorsal large spines
Ch. novenarius: dorsal large spines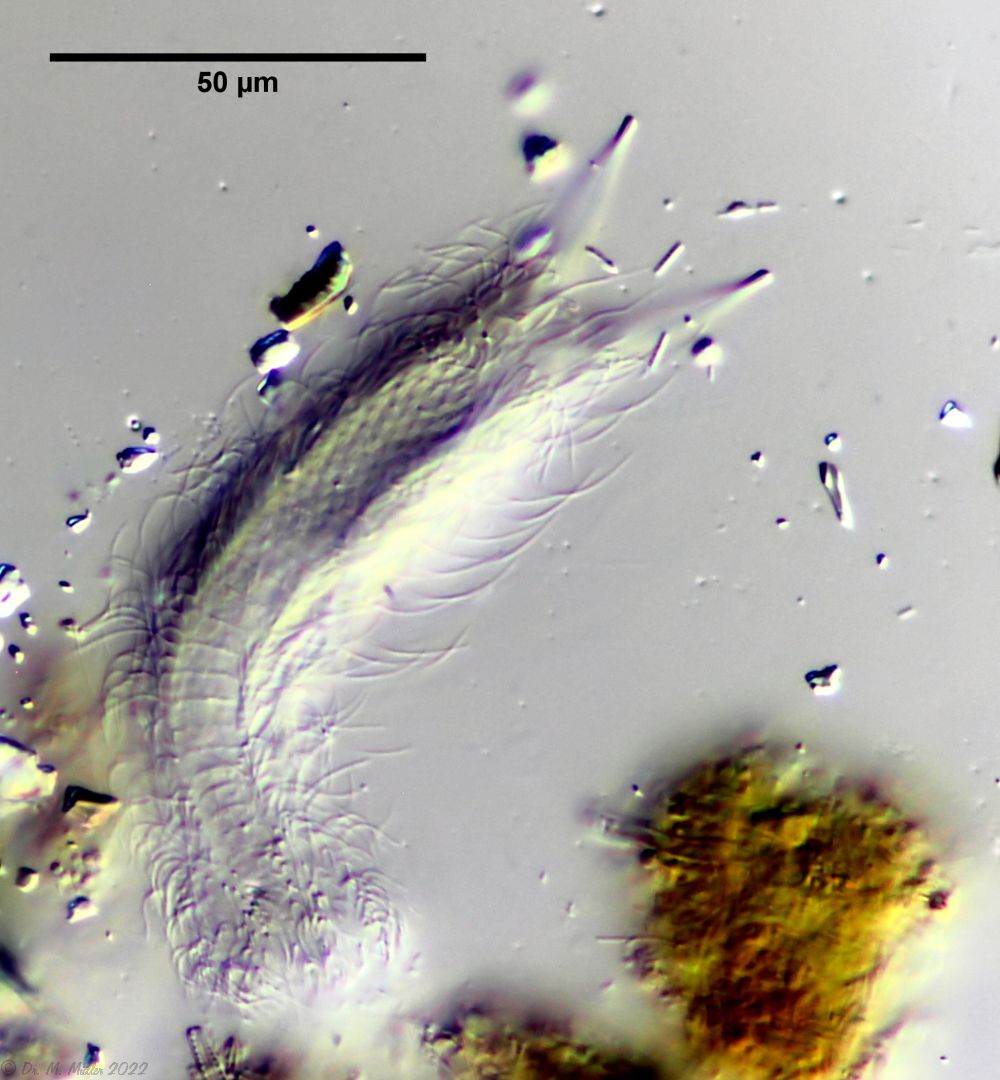 Ch. novenarius: ventral
Ch. novenarius: ventral
 Ch. acanthocephalus: focus on the dorsal scale
Ch. acanthocephalus: focus on the dorsal scale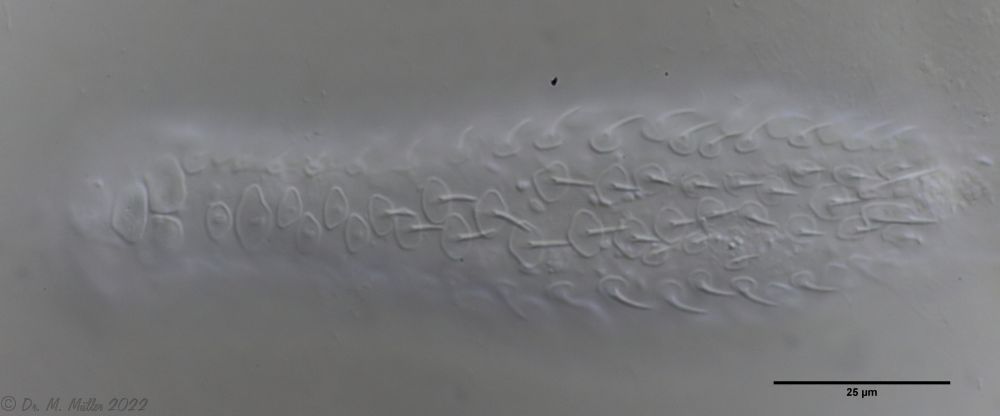 Ch. acanthocephalus: ventral
Ch. acanthocephalus: ventral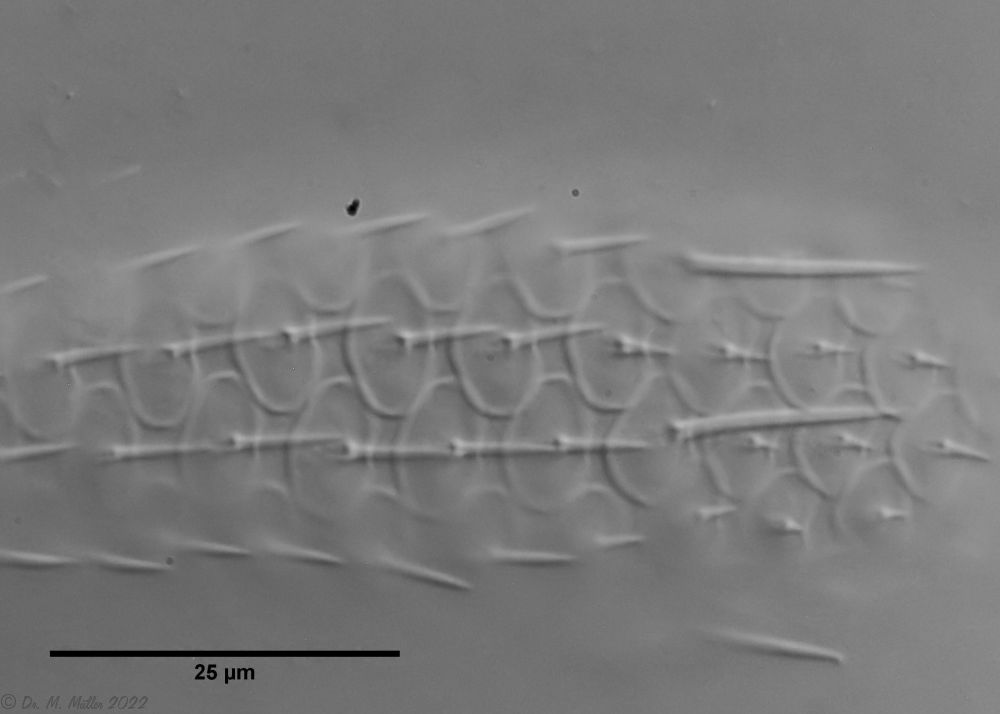 Ch. acanthocephalus: Macerated
Ch. acanthocephalus: Macerated Ch. acanthodes: dorsal scales
Ch. acanthodes: dorsal scales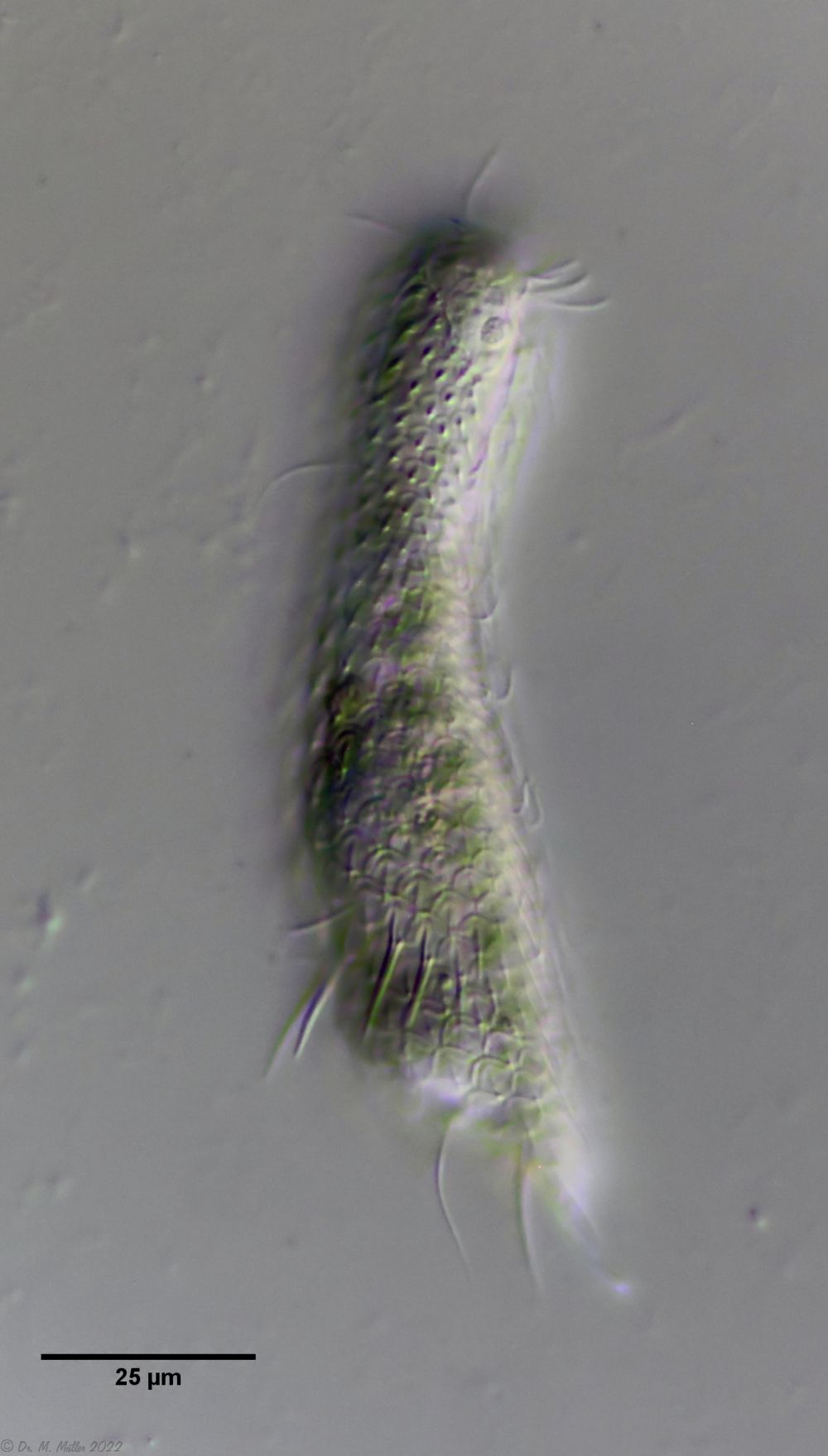 Ch. acanthodes: side view
Ch. acanthodes: side view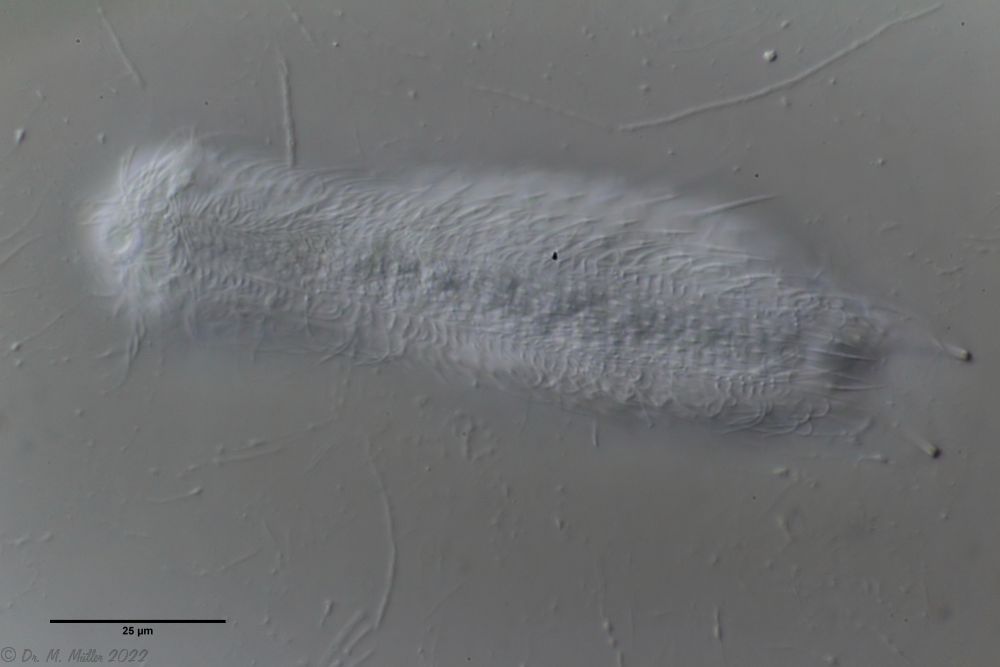 Ch. acanthodes: ventral scales
Ch. acanthodes: ventral scales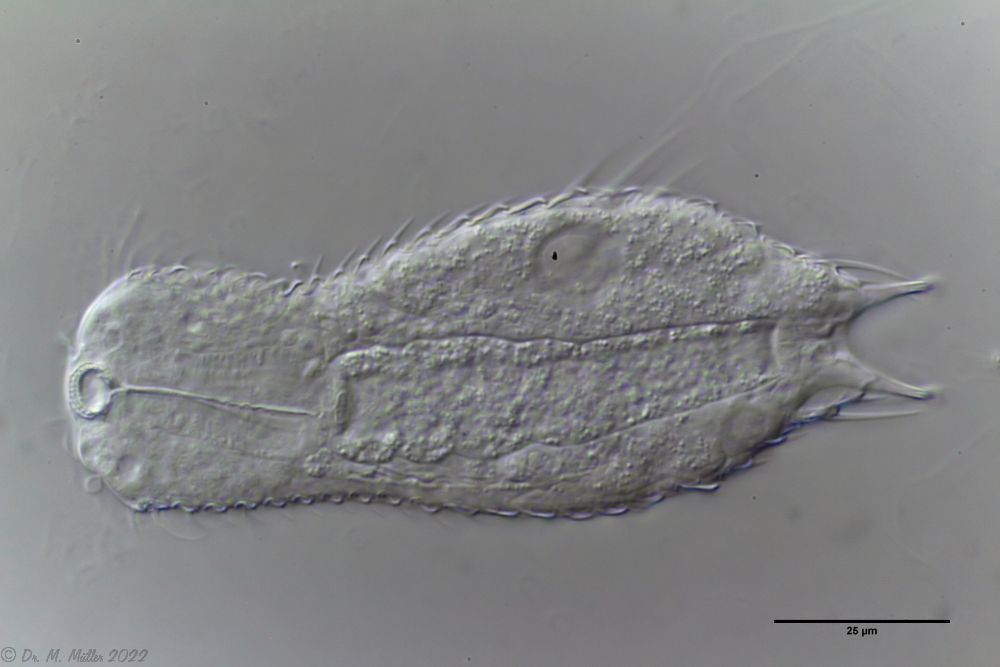 Ch. acanthodes: Cross-section
Ch. acanthodes: Cross-section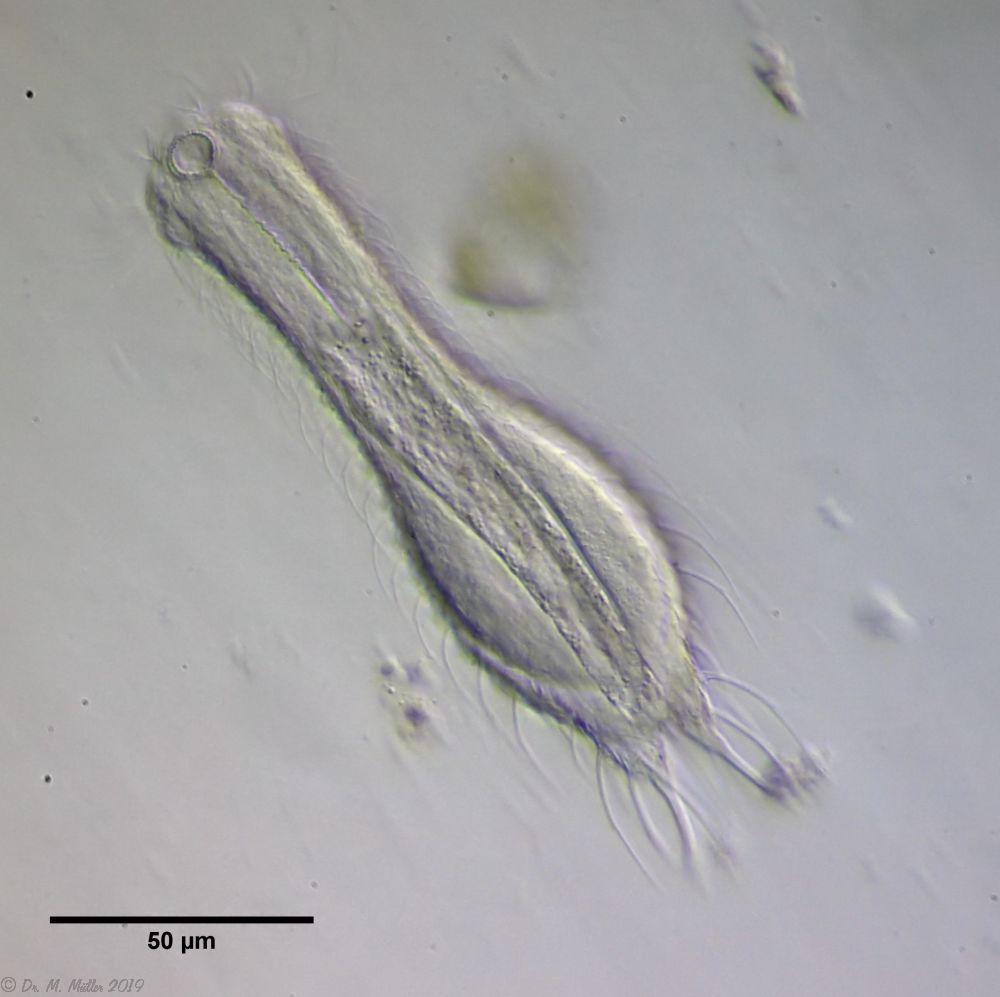 Ch. cordiformis: cross section
Ch. cordiformis: cross section Ch. cordiformis: free swimming specimen with focus on the scale spines
Ch. cordiformis: free swimming specimen with focus on the scale spines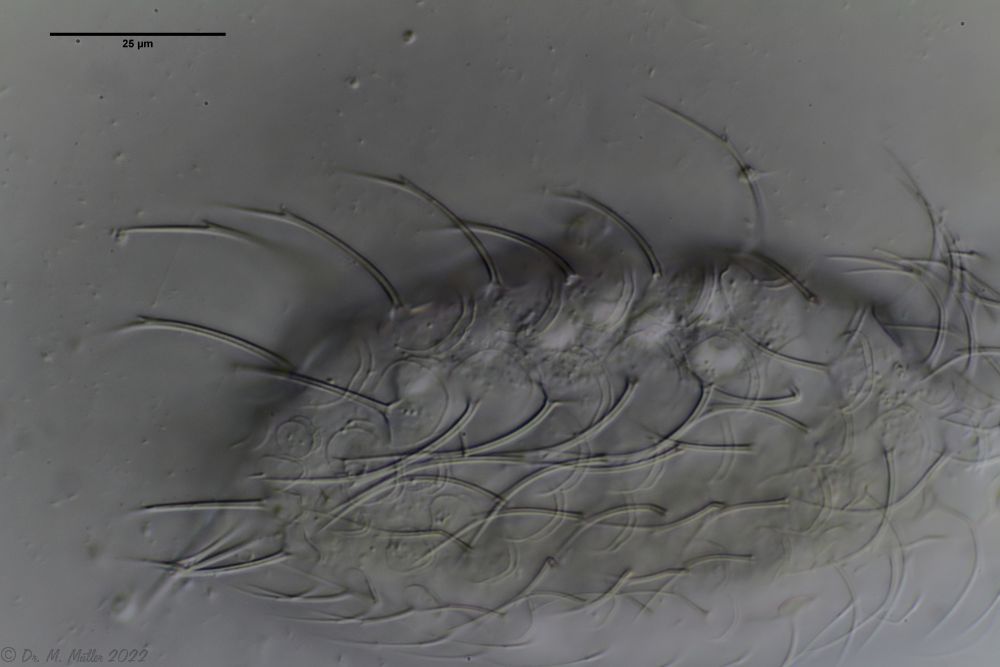 Ch. cordiformis: spiny scales with secondary tip.
Ch. cordiformis: spiny scales with secondary tip.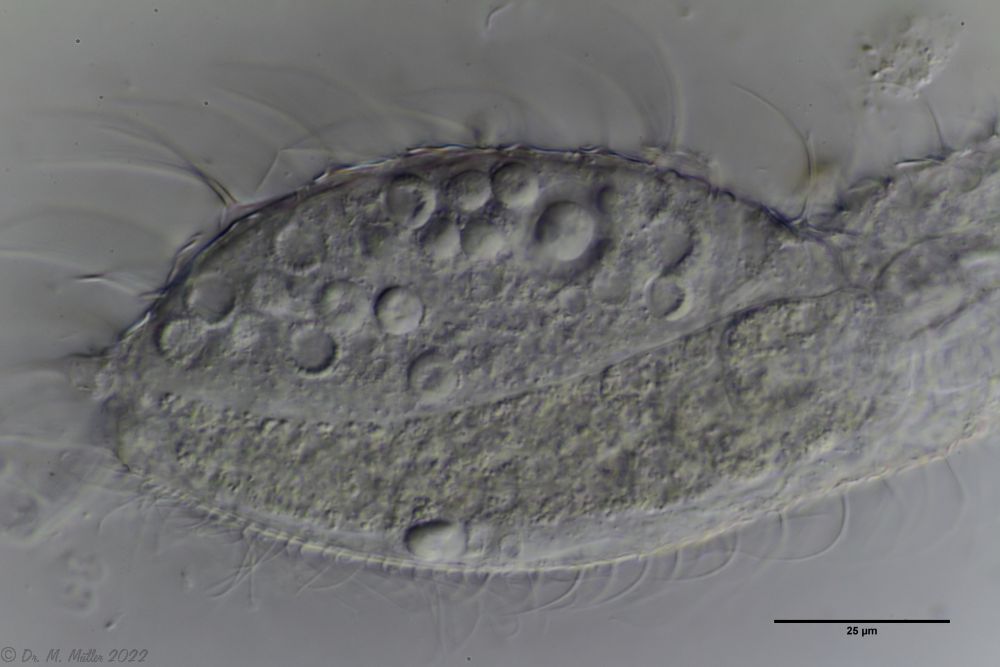 Ch. cordiformis: spherical inclusions.
Ch. cordiformis: spherical inclusions.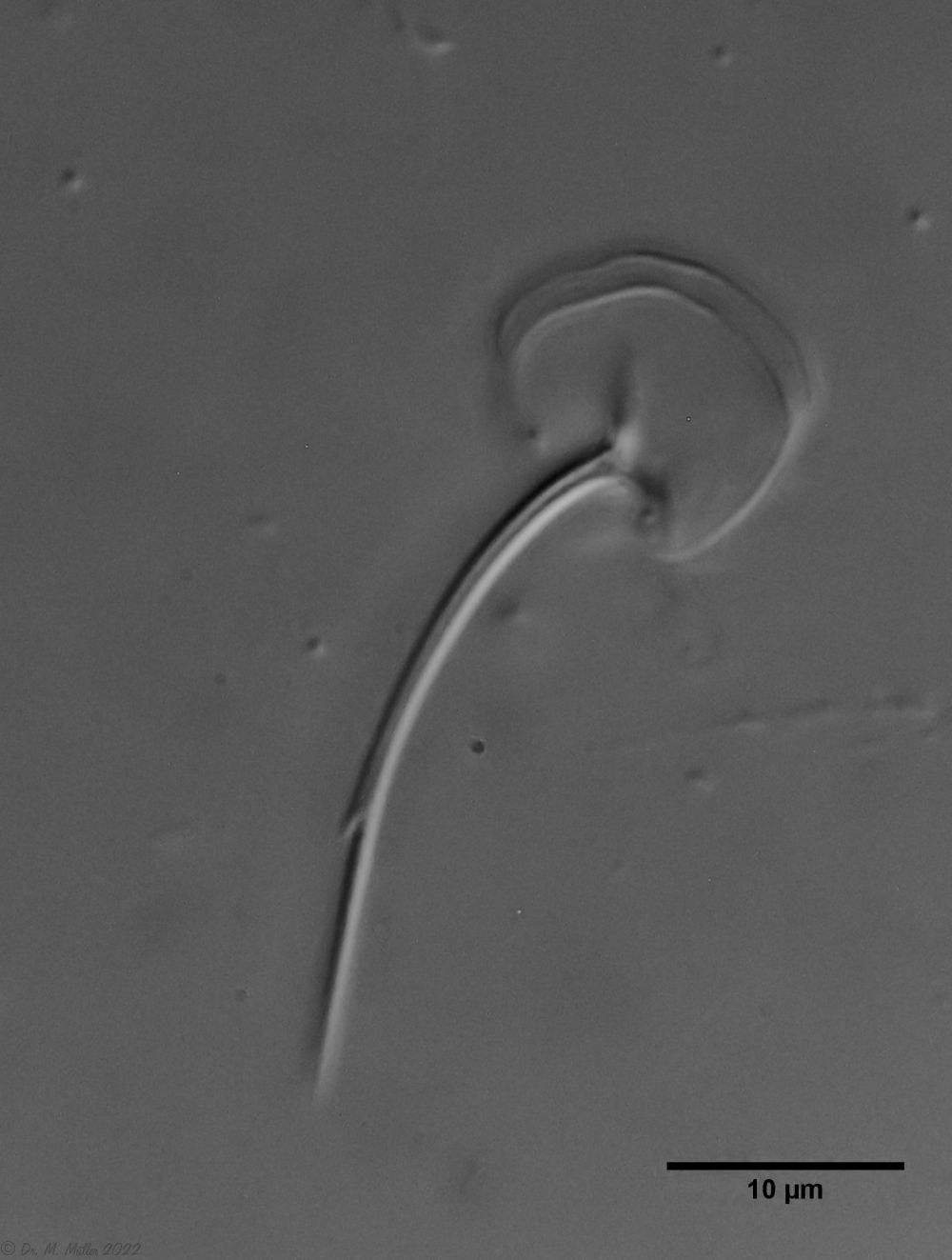 Ch. cordiformis: heart-shaped scales
Ch. cordiformis: heart-shaped scales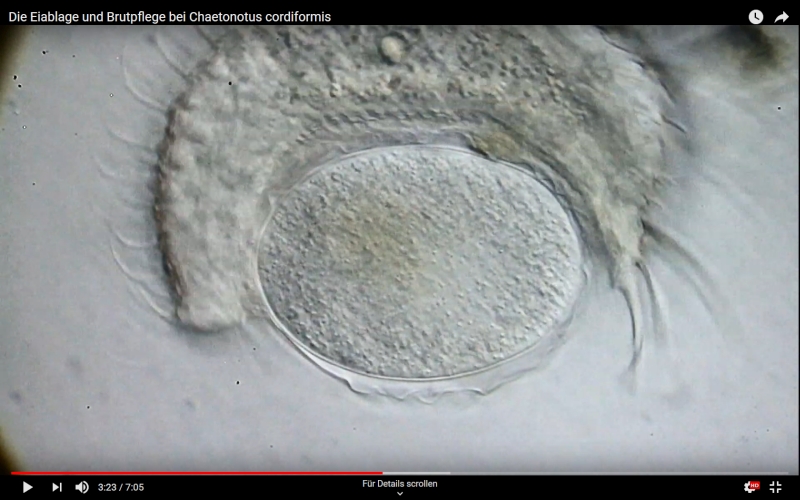
 Dorsal; ds: dorsal spines (5 pairs).
Dorsal; ds: dorsal spines (5 pairs).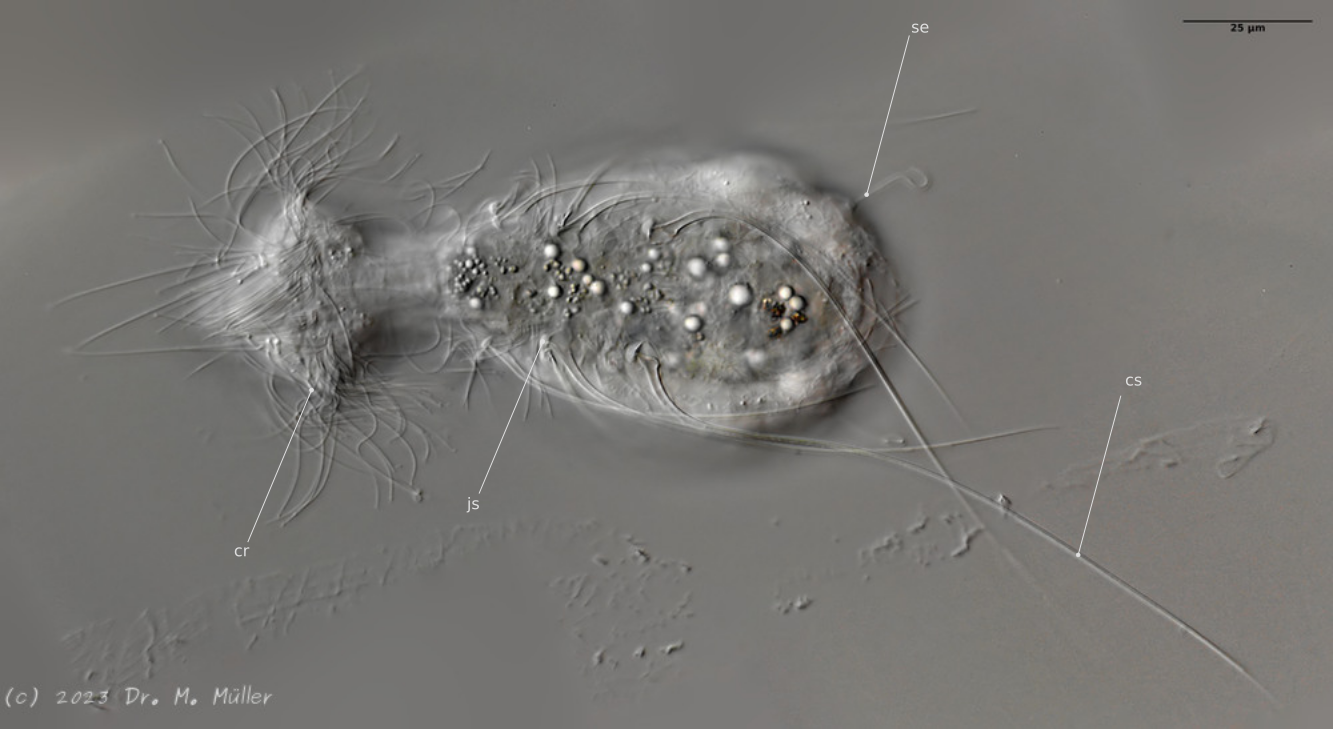
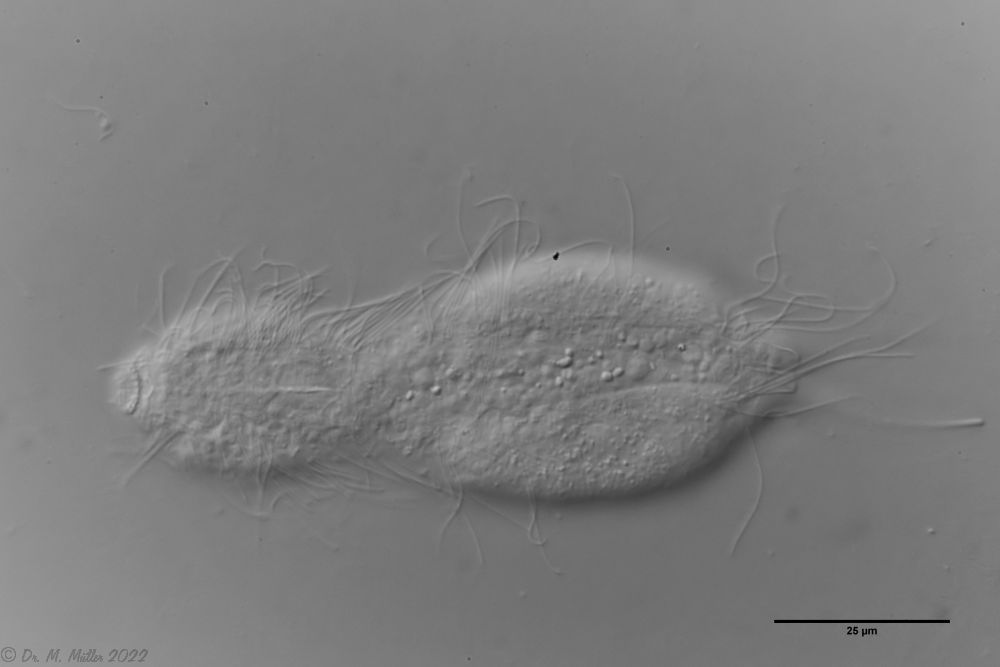
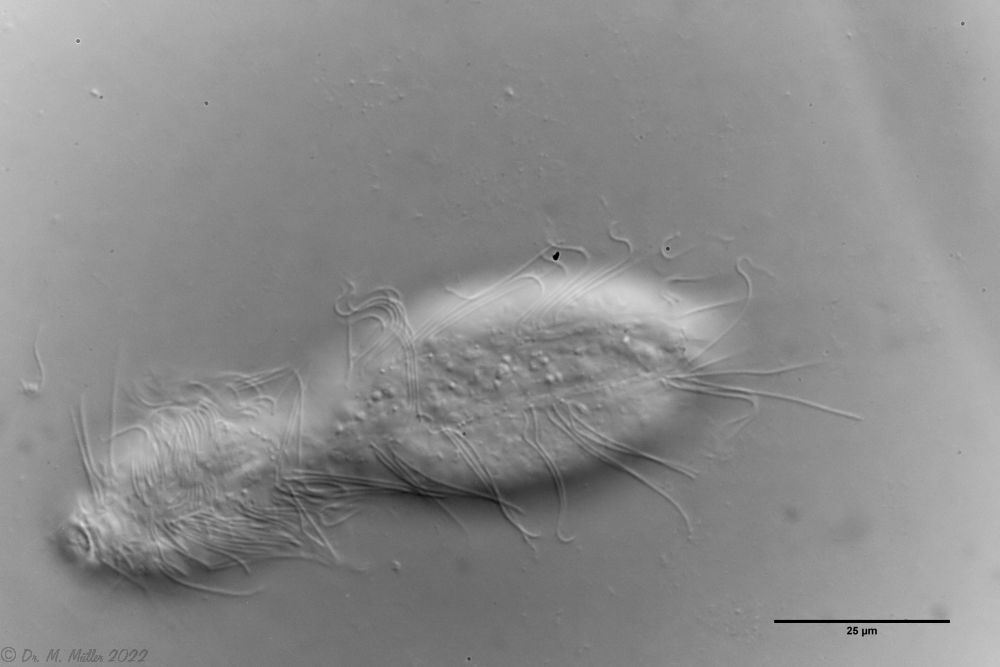
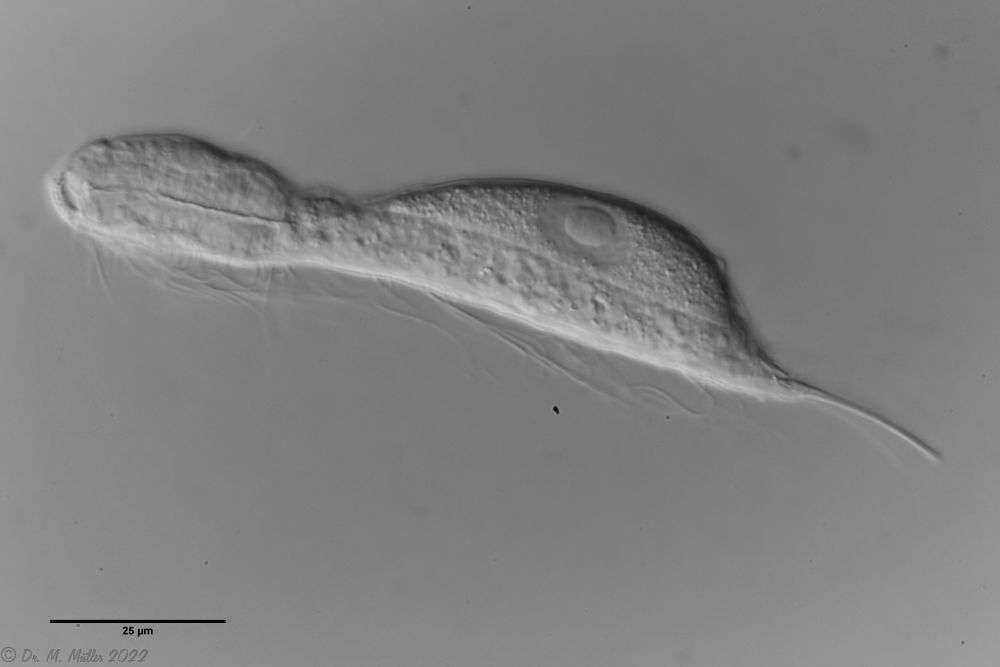
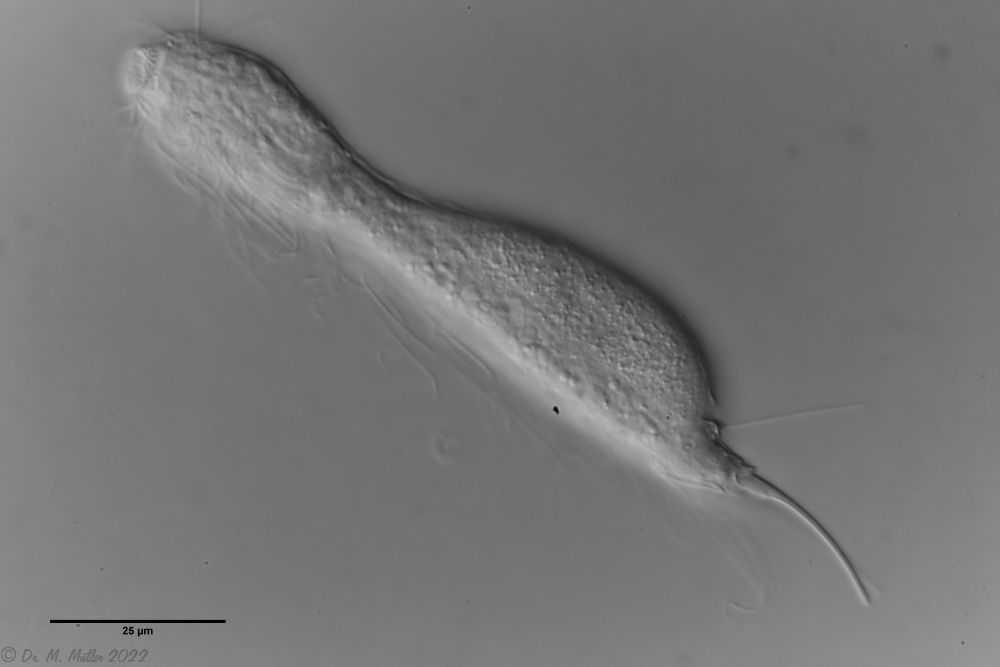
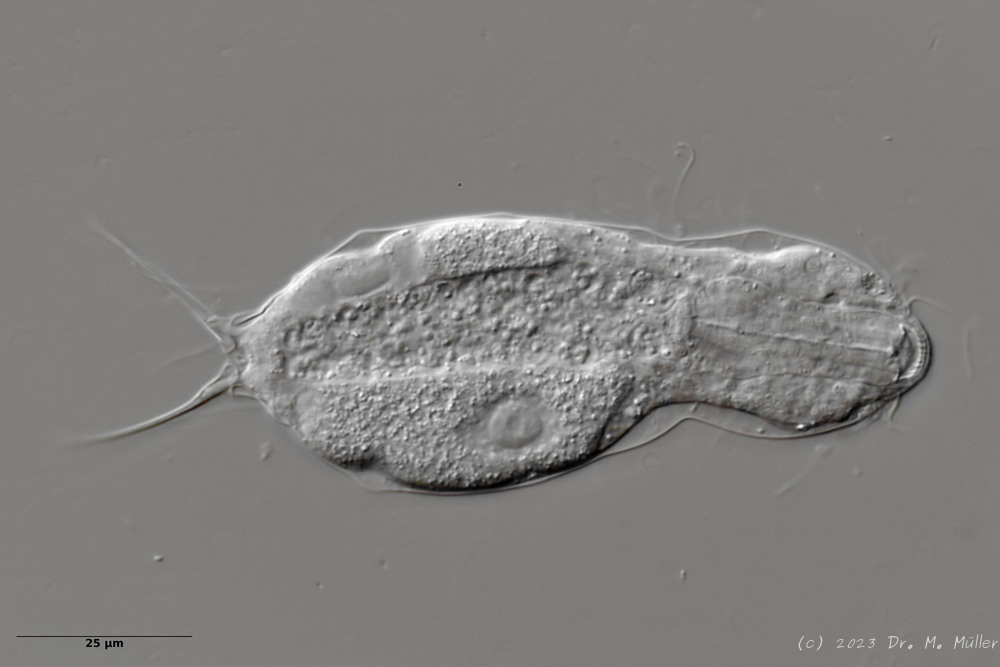
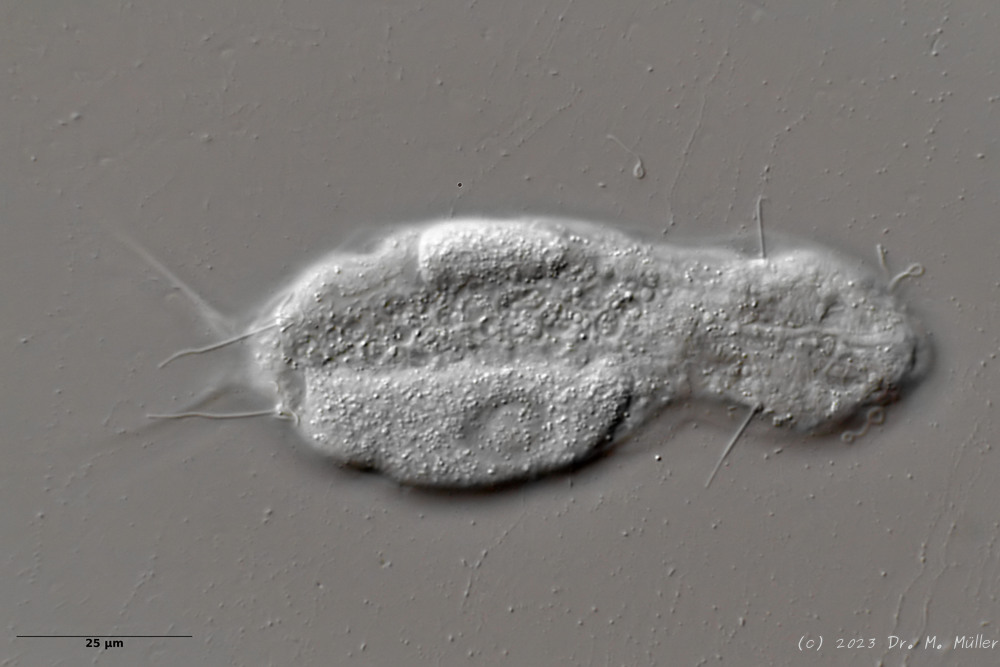
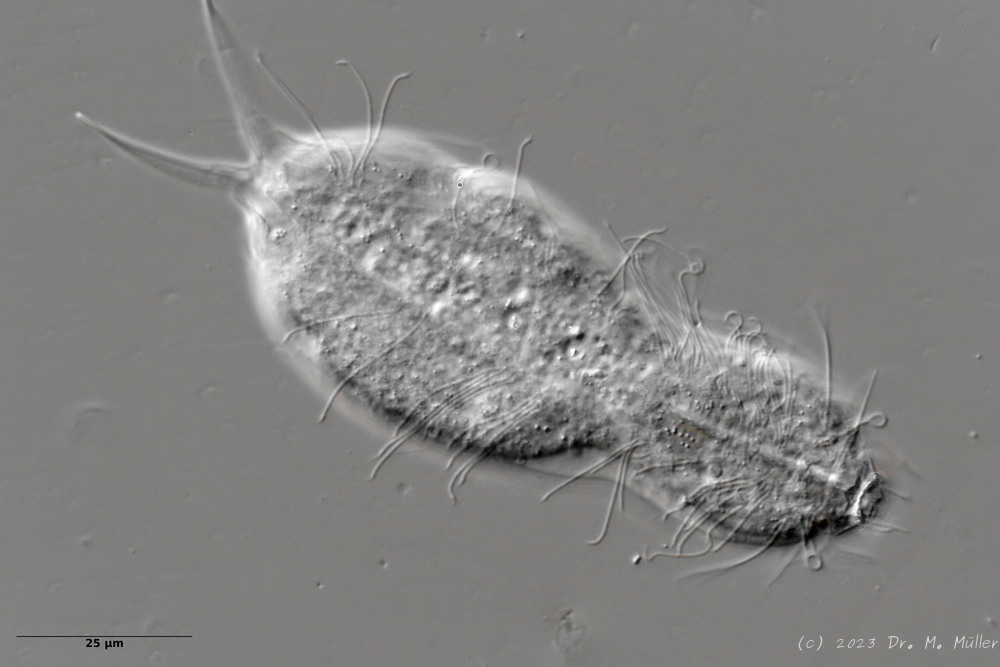
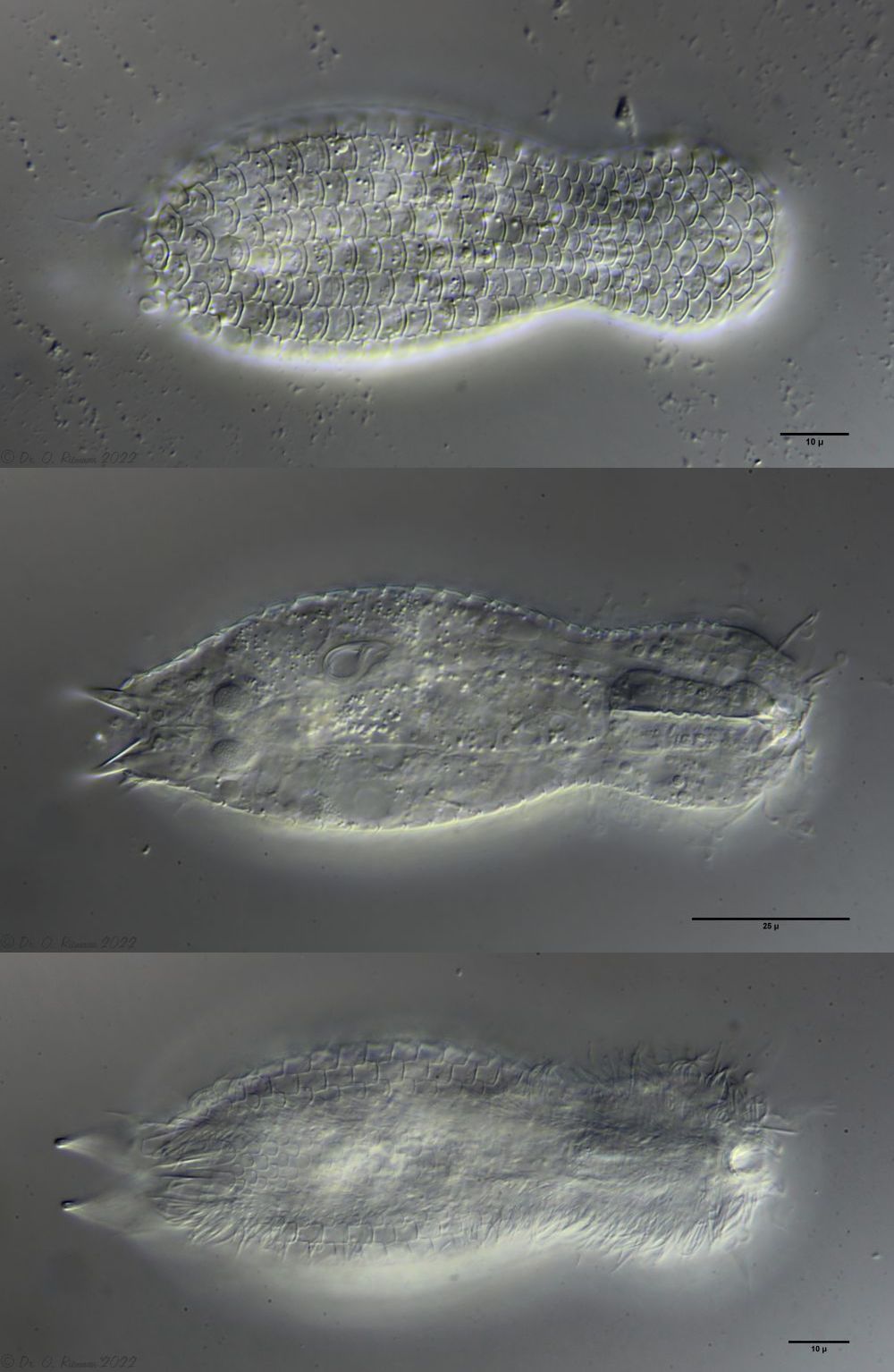 L. minor chaetifer: above dorsally with smooth scales typical of Lepidodermella;
middle in cross section with X-organ and sperm bundle;
below ventral scaling with distinct terminal plates ( Ole Riemann).
L. minor chaetifer: above dorsally with smooth scales typical of Lepidodermella;
middle in cross section with X-organ and sperm bundle;
below ventral scaling with distinct terminal plates ( Ole Riemann).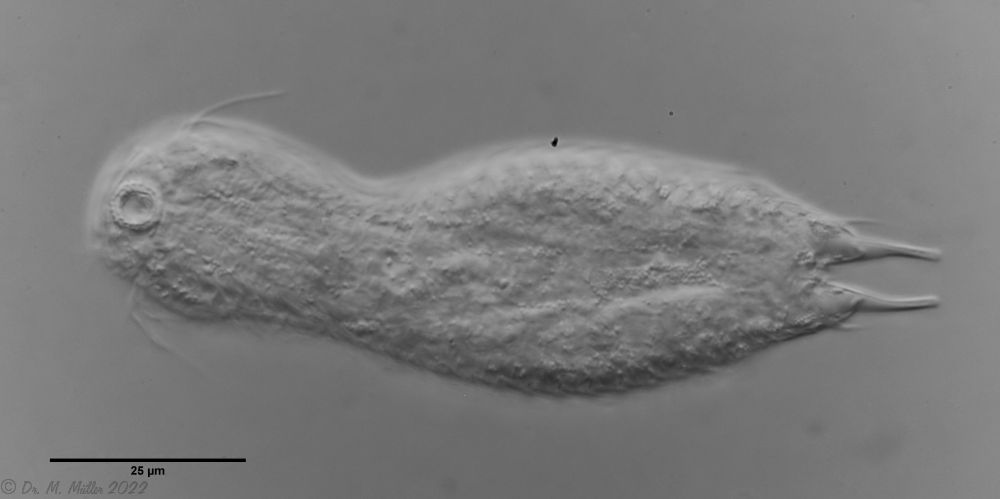 L. minor chaetifer: the pair of spines at the base of the toes is characteristic of the subspecies chaetifer
L. minor chaetifer: the pair of spines at the base of the toes is characteristic of the subspecies chaetifer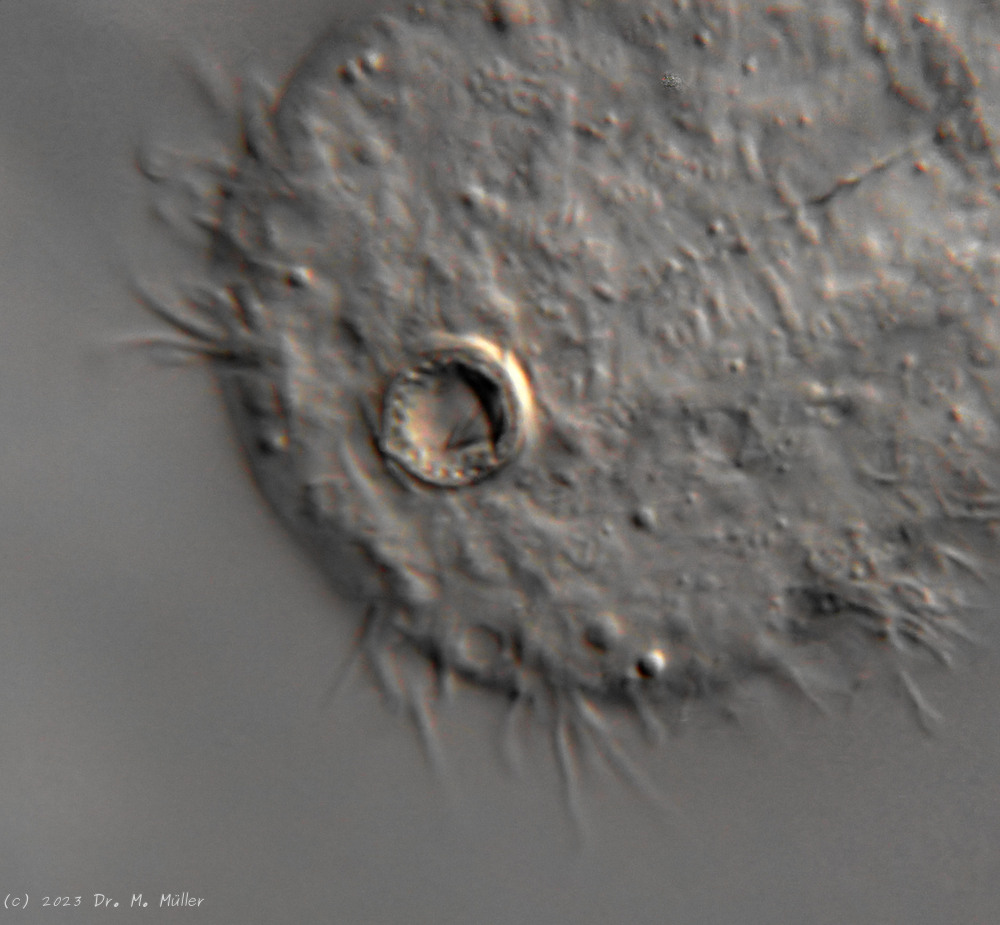
 Dorsal scales
Dorsal scales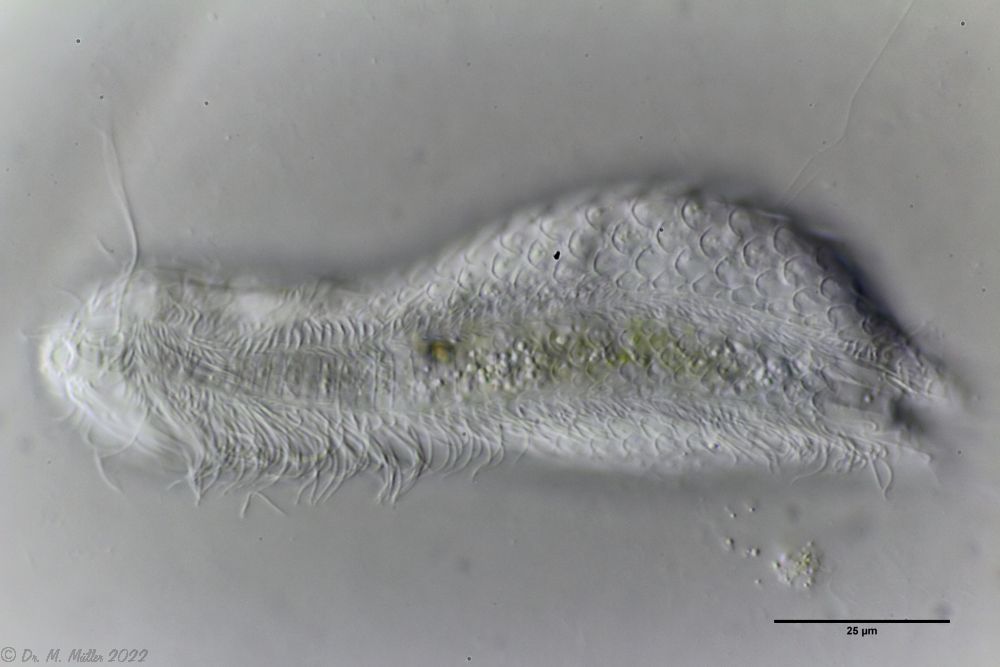 Ventral scales; large terminal plates; rectangular braces under neck
Ventral scales; large terminal plates; rectangular braces under neck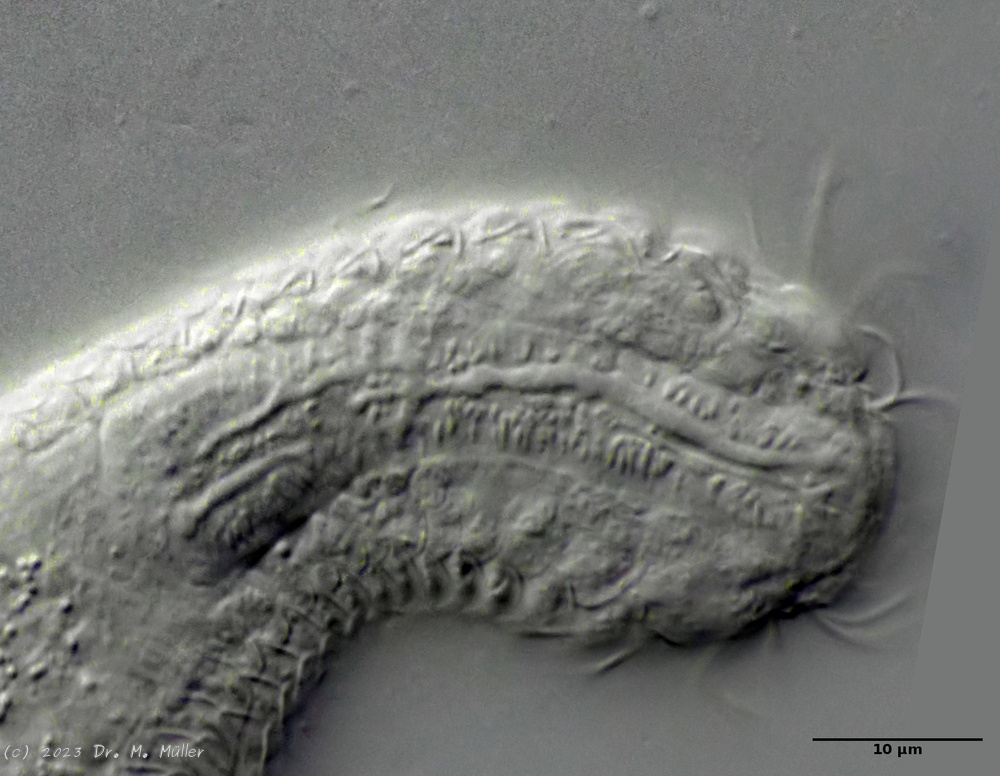

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}