Gastrotrichs - or hairybacks - are microscopic multicellular animals living hidden in almost any water body, unknown to most people. Yet they are the most abundant multicellular organisms in many habitats and, together with rotifers, form the basis of the multicellular fauna in freshwater. Nevertheless, comparatively very little is known about their life cycle and their biology, and many an enthusiast of microscopic study has had no or only a very fleeting encounter with these beautiful and interesting animals, even after years of study of the “life in the pond”.
Therefore, I would like to dedicate a web-site to these shy stars of the pond, where freshwater gastrotrichs will be described in detail. Marine gastrotrichs are not treated on these pages. Therefore - if not explicitly stated otherwise - the term “gastrotrichs” is used exclusively for the freshwater fauna.
This web site is intended to assist both the dedicated amateur and the professional observer in the microscopic observation and identification of freshwater gastrotrichs. Therefore I try to collect short descriptions from the literature for as many known freshwater species as possible and present them in a systematic order. If possible, this information is supplemented by high quality photographs (where appropriate, photographically documented species are marked with a “:camera:”) and identification hints for light microscopic species diagnosis are given, which have proven successful in my observations.
Statistical Information
- Of the 397 freshwater species described, this web site includes for 379 Species individual pages.
- Of these species, no literature references could be found for 324 species
- For the remaining 42 species a profile from the literature was taken over in each case
- Observation photos were added to these profiles for 0 species observation photos were added
The second part deals with the biology of freshwater gastrotrichs. In addition to the anatomy of the hairybacks, their peculiar life cycle is also discussed here.
By using microaquariums, it was possible to observe hairybacks continuously over an extended period of time (several weeks). This provided unique information about the behavior and development of gastrotrichs.
Statistical Information
- In total this web site contains 0 individual pages on the topic "Biology"
Finally, in the section “Technique” hints shall be given on the necessary or by me used apparative equipment as well as on the preparation techniques used for the observation of hairybacks. Here you will find instructions that allow, for example, high-resolution observations or to perform long-term studies of the development and behavior of gastrotrichs.
Statistical Information
- In total this web site contains 9 individual pages on the topic "Technique"
As with all web sites, this one is never finished and competes with my other interests for my scarce time. Therefore - despite all care and dedication - there will always be gaps and mistakes in the content. The gaps I hope to close in the course of time - with the errors I depend on your assistance. Please don’t hesitate to point out problems or inaccuracies - I’m always grateful for constructive feedback!
This web-site does not consider itself a scientific publication. Therefore, in favor of clarity and simplicity, I refrain from the last “scientific” caution on some topics. For example, in order to achieve the necessary scientific certainty in species identification, it will often be necessary to verify the species diagnosis using the given literature. This web site is only intended to facilitate a first introduction to the great subject of “gastrotrichs”.
Subsections of The Small World of Freshwater Gastrotriches
Naturbelassenen Gartenteich; Einträge von Proben aus verschiedenen Standorten
Mürmes
Moor in verschiedenen Übergangsstadien, Schwingrasen auf abgetorften Bereichen, Feuchtwiesen mit Fließgewässer
Dr. Stephan Krall
Purren
Der Teich bei dem Hügel “Purren” wurde von Martin Kreutz ausgiebig beprobt und untersucht (:up_arrow: Beschreibung von M. Kreutz). Netterweise hat er mir Proben von diesem reichen Fundort zu Verfügung gestellt und mir so die Möglichkeit eröffnet, die reiche Gastrotrichenfauna dieses Waldtümpels zu untersuchen.
Dr. Martin Kreutz
Ellenbachweiher
Hangarsteinsee
Am Hangarsteinsee
Weichselbrunner Weiher
Sima Moor
Simmelried
Das “Simmelried” (:up_arrow: Beschreibung von M. Kreutz), ein Feuchtgebiet in der Nähe von Konstanz, wurde jahrelang systematisch von Martin Kreutz untersucht. Netterweise hat er mir Proben dieses “biologischen Hotspots” zu Verfügung gestellt und mir dadurch ermöglicht, die Gastrotrichenfauna dieses Gebietes näher zu untersuchen.
Dr. Martin Kreutz
Gartenteich
Privacy Policy
Your data is important to me! That’s why I created this web site with static individual pages that do not store any data from the visitor either on the server side nor on the visitor side (as cookies). Only the data technically necessary for communication is stored temporarily.
I do not use any third-party products on this website where I could not ensure that your data is handled responsibly. In particular, I refrain from using products from Google (Alpha) and Meta (Facebook), whose business model is aimed at marketing your data.
While this waiver is painful at times, I believe it is the only way to treat your data fairly. For this reason, I do not embed YouTube videos directly on my website. A click on the corresponding “fake player” pictures leads you directly to the pages of Youtube and shifts the data protection problem from me to Youtube…
I incorporate the maps of the service “OpenStreetMap” (https://www.openstreetmap.org), which are offered on the basis of the Open Data Commons Open Database License (ODbL) by the OpenStreetMap Foundation (OSMF). OSMF Privacy Policy.
To the best of my knowledge, user data is used by OpenStreetMap solely for the purposes of displaying the map functions and caching the selected settings. This data may include, in particular, IP addresses and location data of users, which, however, are not collected without their consent (usually executed in the context of the settings of their mobile devices).
Pharyx ( two bulbi without intermediate piece ): 15 µm - 16 µm
Diameter of the mouth ( narrow, extended, terminal ): unknown
Dorsal scales: 12 - 16 thick, curved, simple and immobile spines; of which 3 dorsally (29-30 µm), 9-13 laterally, extending around the posterior end(18-21 µm)
Ventral scales: two ciliated bands, united anteriorly, visible dorsally and laterally
Diameter of the mouth ( elliptical, two teeth (9x5 µm) ): 8 x 20 µm
Dorsal scales: no scales, but closed cuticular layer
Ventral scales: broad hypostomium with transverse furrow (38-42 µm); two rows of cilia; additionally two ciliary tufts on head (median), one pair of ciliary tufts laterally on head
Oecology: Psammon (sandy ground)
Similar species: Similar to Ichthydium, but completely covered with structureless cuticle (in Ichthydium the cuticle is usually finely structured); well demarcated by complex structure of mouth.
The genus Aspidiophorus is very easily recognized microscopically by the apparent “double contour” of the body. This double contour is produced by the typical style scales of this genus, which consist of a base plate that sits directly on the cuticle. From this base plate rises a thin peduncle, which carries at its end a - usually larger - un-spined terminal plate:
typical scales of an Aspidiophorus, here A. squamulosus.
This results in the typical double outer contour of the animals:
Diagnosis
Body bottle shaped
Furka normal
Peduncle scales
Subsections of Aspidiophorus
Aspidiophorus aster
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Aspidiophorus Voigt, 1903
Species: aster
Length ( bottle ):
Width:
Width of the head ( five-lobed ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( cylindrical ):
Diameter of the mouth ( around ): unknown
Unfortunately, I have no data available for this species!
If you have access to the following literature:
Martin,L.V. 1981. Gastrotrichs found in Surrey. Microscopy 34: 286-300.
I would be very happy if you would send me a copy by Mail!
Unfortunately, I have no data available for this species!
If you have access to the following literature:
Grosso,L.E. 1973. Notas sobre Gastrotrichos Argentinos II. Neotropica 19(59): 87-89.
Grosso,L.E. 1973. Notas sobre Gastrotrichos Argentinos I. Chaetonotus (Zonochaeta) guruguetoi sp. nov. y Aspidiophorus brahmsi sp. nov. Physis 32B: 133-137.
I would be very happy if you would send me a copy by Mail!
Width of the head ( three lobed ): 17 µm - 18.5 µm
µm
Length of the furca: 20 µm
Adhessive tubes: 50% of furca
Pharyx: 32 µm - 36 µm
Diameter of the mouth ( around ): 5.5 - 8.5 µm
Dorsal scales: rhombic petiole scales (3µm long), 41-42 per row, last row in middle with spines (9-14µm), laterally at base of toes a pair of long spines (16-18µm), sometimes with another pair of spines or elongated scales
Ventral scales: Interciliary field with a pair of terminal spines; very many minute keels
Unfortunately, I have no data available for this species!
If you have access to the following literature:
Grosso,L.E. and Drahg,F. 1983. Gastrotricos dulceacuicolas de la provincia de Tucman. I. Chaetonotus soberanus sp. nov. y Aspidiophorus lilloensis sp. nov. Neotropica 29: 189-193.
I would be very happy if you would send me a copy by Mail!
Pharyx ( cylindrical, with small terminal swellings ): 30 µm - 35 µm
Diameter of the mouth ( around ): 3 µm
Dorsal scales: 9 rows of 16-17 peduncle scales each; end plates (6-7 x 2-3 µm) rounded anteriorly, incised posteriorly, no keels; last 2-3 rows terminate in 20 µm spines; lateral on toes a pair of spines (25 µm).
Ventral scales: Ventral intercilliary field with 25 rows of tiny petiole scales
Oecology: Mud dweller
Similar species: A. longichaetus
: Head rather trilobate, no median spines
Dorsal scales: 41-44 rows of 48-50 small peduncle scales each; end plates elliptical, acuminate with keel, overlapping, 3 x 1.5 µm; two 5x2 µm peduncle scales on furca inner side.
Ventral scales: Ventral innercilliary field with two terminal plates (7x4 µm) and 10 rows of narrow, distally pointed keel scales (4x0.75µm)
Dorsal scales: 25 rows of 40-45 rhombic petiolar scales each with lateral and median keels (4x6 µm); petioles 6-7 µm; terminal plates of last terminal rows often enlarged (12-17 µm); posterior end free; between toes 4 short spines on oval scales.
Ventral scales: Ventral innerciliary field covered with petiole scales
Oecology: Sludge dweller
Similar species: distinctive, variable type
Particularities: 3 teeth, eggs smooth (125x74 µm)
Aspidiophorus paradoxus is the largest Aspidiophorus species with about 300µm length.
Dorsal scales
The entire animal is covered with relatively large rhombic peduncle scales.
Cross section
In cross-section, the structure of the petiole scales is clearly visible: the scales sit with a small base plate on the cuticle of the animals, from which rises a thin, hollow pedicel. At the end of the peduncle sits a rhombic terminal plate, with a central keel. At the posterior end of the animal, the terminal plates of the last row of scales are enlarged.
The pharynx of the animal is terminally swollen, and the head is weakly five-lobed with two separate pairs of palpal tufts.
Central view
Ventrally, the strong hypostomion behind the mouth opening is striking. The two ciliated bands split at the head, but the inner branches do not unite in the population I examined. The base of the toes does not bear scales, the adhesive tubes
measure about 50% to 70% of the toe length and taper to a point.
Let’s take a closer look at the scales:
Backscales
In the scales on the back, the rhombic shape of the end plates is most clearly recognizable. Less conspicuous - but typical for the species - is the central keel of the scales.
Cross section shed
In cross-section, the complex geometry of the stem scales becomes clear - base plate, stem and end plate form a very flexible and stable armor. The additional cavity under the outer scales acts like a “crumple zone” and further increases the protective effect.
Cross section scale stems
The stems of the scales consist of hollow tubes that ensure maximum stability with minimum material input - a fascinating example of evolutionary optimization.
The abdomen of the animals is not completely covered with petiolar scales. Rather, they end in the anal region and are joined by simple small, rounded keel scales that are not an obstruction to feces.
Scaling of the abdomen.
At the base of the furca some (according to literature 4) spines protrude into the toe.
The head is almost completely covered with slightly smaller peduncle scales:
Headshed
Cephalion and pleurae are quite small and inconspicuous.
According to literature A. paradoxus has three teeth in the pharynx:
Mouth armament
In the animals I examined, there was only one curved stylet brace, the tip of which protrudes into the lumen of the pharynx and is probably used to open algal cells that are conveyed past it. It is possible that the literature reference to “three teeth” is merely based on a microscopic artifact, as the entire clasp may not be in the focal plane as a whole.
Dorsal scales: 45-53 rows of 29-32 petiole scales each (5-7 µm); no keel, not rhombic, similar shape to that of
P. rhomboides
; Toe base dorsal with keels
Ventral scales: 6 keeled terminal plates; at the posterior end 10-12 rows of keels (4-4.5 µm), otherwise naked
Oecology: between roots; Brazil
Particularities: very small pharynx; body very broad
Dorsal scales: 18-20 rows of 42-45 small petiolar scales each; petioles 1.5 µm; terminal plates 2-3.5 µm long, proximally strengthened margin, distally acuminate, weak median keel; no spines
Ventral scales: Ventral intercilliary field naked, except for a few elongate peduncle scales; two narrow terminal plates(6.5-11.5 µm)
Oecology: Bog, mud
Similar species: delimited by naked ventral intercilliary field
Particularities: without pseudocells; naked ventral intercilliary field .
I have found this belly-harper only once so far in a bog (Sima bog). By the naked ventral intermediate field with long terminal plates the species is well to delimitate:
The round anterior margins of the end plates of the lateral peduncle scales are also well visible. The found animal is with ca. 130 µm a little smaller than the literature specification and all other measurements scale accordingly. The mouth ring is very small and strongly marked. A hypostomium does not exist.
In the median optical section the dumbbell-shaped pharynx can be seen well. The scales are distally long extended (unfortunately the shape of the end plates could not be seen). The granular structure of the two adhesive glands is also interesting.
Width of the head ( five-lobed, pointed cephalion ): ? µm
µm
Length of the furca: 16 µm
Adhessive tubes (tapered, claws): 50% of furca
Pharyx ( cylindrical with slight swellings ): 37 µm
Diameter of the mouth ( around ): 7 µm
Dorsal scales: 53 petiole scales (3 µm) per row; 3 pairs of spines at outer base of toes (20-22 µm).
Ventral scales: Ventral intercilliary field very many petiole scales; a pair of terminal spines (8-10 µm) and many shorter spines protruding into the toe cutout.
Oecology: Mud dweller
Similar species: well demarcated by claws and spines
Dorsal scales: 25-30 rows, each with 60-80 minute petiolar scales; terminal plates unkeeled (0.5-2 µm), elongate, pointed distally; posterior end with elongate keel scales
Ventral scales: Ventral intercilliary field numerous longitudinal rows of minute petiolar scales; two keeled terminal plates (6-7 µm).
Dorsal scales: 12-15 rows of 40-42 petiolar scales each with rhombic terminal plates without median keel (3-4 x 5-6 µm); scale shell ends in anal region; posterior end and toes naked; 3 pairs of short spines at base of toes (8-10 µm).
Ventral scales: 2 terminal keels (5-6 µm), 12 rows of keels without base plates
Oecology: Mud dweller
Similar species: A. paradoxus
: unkeeled basal plates; spination at the posterior end; mouth grafting
Of the (acc. (Schwank, 1990))
twenty limnic Aspidiophorus species only four are listed in the “official” species lists for Germany. Another species was described some time ago by Michael Plewka. The species Aspidiophorus squamulosus has been found so far only in Poland and in western France - so it is no wonder that this animal is also native to us, between the previous finding areas.
Fig. 1: Aspidiophorus squamulosus; optical section and scale image.
The internal structure of the animal becomes quite clear in the side view:
Fig. 2: A. squamulosus, side view.
A. squamulosus is with ca. 230µm somewhat smaller than the similar species A. paradoxus and is clearly distinguished by the three spines each above the toes (pic. 3, ob. li. and mi. r.). In addition, the terminal plates of the petiolar scales do not possess a median ridge. Also the prominent, furrowed hypostomium (Fig. 3, ob. r.) is typical for this species. Some animals I found were in their “hermaphroditic phase” (post-parthenogenetic phase) and showed a very large X-organ and sperm bundle (Fig. 3, mi. l.). One animal also carried a very large egg (with a pronounced nucleolus), which it laid during the approximately 10-day observation period. Unfortunately, I was not able to observe the oviposition itself (also the egg was not found in the specimen).
Figure 3: A. squamulosus: details;
up. l.: toe spines; up. r.: hypostomium
mi. l.: X-organ and spermathecae (marking); mi. r.: toe spines and toe scales;
low. l.: transverse section of scale carapace; low. r.: stylet (mark)_
As is often the case, there were some deviations from the species description in the literature. With up to 50 generations per year and years of isolation in a small garden pond, I find minor deviations not surprising. However, it is possible that some details were not noticed in the rare, earlier observations. For example, the base of the toes should be unscaled (Fig. 3, mi. r.). However, the specimens I observed show many very small, roundish and close-scaled scales. Only the conspicuous peduncle scales do not appear at the posterior end of the animal.
In some Aspidiophorus species movable “teeth” in the mouth tube are described. Although A. squamulosus should be toothless, two actively movable styletes could be observed, the tip of which ended in the mouth tube and reminded of tardigrades (Fig. 3, un. r.).
The peculiar stipe scales of the genus Aspidiophorus are not to be grasped in their three-dimensional beauty in the scale association. Therefore, I subjected a pitiable specimen of A. squamulosus to scale analysis:
Fig. 4: A. squamulosus; colored and - unfortunately - flattened single scales.
Since during a scale analysis just the very small scales are pressed rather flat, I tried a drawing reconstruction of the three-dimensional structure:
Fig. 5: A. squamulosus; bottom: composite scale image; top left: isolated and stained scales; top right: drawing reconstruction of a single peduncle scale.
Another point that has remained open so far is the diet of A. squamulosus. The main diet consists of relatively large unicellular algae or eye flagellates. To swallow this large prey, the mouth of the animals is equipped with movable lamellae that can be folded outward for feeding (cf. Captochaetus). This allows the diameter of the mouth opening to be greatly expanded, unlike most gastrotrichs. During the swallowing movement, the stylet is pressed into the mouth tube and the prey cells are guided past it. In the process, the hard cell wall is probably perforated so that the digestive secretions in the intestine can penetrate the cells.
Fig. 6: A. squamulosus; intestine filled with ocular flagellates.
If one examines the mud of stagnant waters microscopically, one always finds gastrotrich clutches in empty shells of water fleas with sometimes dozens of eggs.
Fig. 7: A. squamulosus; focal sac of a gastrotrich clutch containing eggs of at least three different species.
Often these clutches consist of eggs of different species. Apparently, a number of species prefer to lay their eggs in water flea shells where other gastrotrichs have already laid their eggs. In such a clutch often relatively large (95µm x 55 µm), spiny eggs are noticeable, which have not yet started embryonic development and probably represent permanent eggs.
Fig. 5: A. squamulosus; permanent egg.
One egg of the shown clutch started to develop and turned out to be an egg of the species A. sqamulosus. Therefore I can show here the eggs belonging to the species.
Dorsal scales: 15-20 rows of 38-40 very small peduncle scales each; endplates very delicate and unkeeled (2.5-3 µm); strongly bent peduncles -> scales protrude like bristles
Ventral scales: Ventral intercilliary field 9-10 rows keels; anteriorly mostly naked; no terminal plates.
Oecology: creek
Similar species: well demarcated by protruding scales
Particularities: protruding scales; body cylindrical
Dorsal scales: 15-17 rows of 33-40 petiole scales each; end plates unkeeled 2-5µm, rounded distally and proximally; base of toes naked; 2 pairs of thick spines (8-10µm and 4-5µm) on oval scales with two keels.
Ventral scales: 2 terminal keels (7.5-11µm); in gut region 8-9 rows of tiny keels, otherwise naked
Oecology: montane lake
Similar species: delimitable by spines
Particularities: 2 pairs of spines at the rear end
A. tetrachaetus is a very rare gastrotrich, so far confirmed only for Poland.
A. tetrachaetus: dorsal ((c) Ole Riemann)
The dorsum of the animals is covered with relatively large, unkeeled and rounded pedunculate scales, which do not continue to the base of the toes at the posterior end. A pair of large spines is conspicuous at the posterior end of the scale coat.
A. tetrachaetus: t.s. ( Ole Riemann)
In the cross section the structure of the petiole scales can be seen well.
A. tetrachaetus: ventral (copyright: Ole Riemann)
The ventral intermediate field of the animals is covered only in the intestinal region by 8-9 longitudinal rows of small keels. The pharyngeal region is naked. Two terminal keels form the posterior shoot of the ventral scaling.
A. tetrachaetus: two pairs of spines at the hind end.
Only a closer look at the dorsal toe base of the animals reveals two additional, smaller spines.
A. tetrachaetus: single tooth in oral cavity.
Contrary to the description in the literature, a single tooth can be seen protruding asymmetrically from the oral cavity in the animals at Sima Moor.
Adhessive tubes (three-part (17, 10, 7 µm)): 80% of furca
Pharyx ( dumbbell-shaped ):
Diameter of the mouth ( around ): unknown
Dorsal scales: 7-9 rows of 17-18 unspined scales each; anterior 2/3 round to hexagonal, last 5 rows 3-5 larger scales each.
Ventral scales: Hypostomium large, Ventral intercilliary field with 3-5 rows of 15-17 spineless, rectangular scales each; 2 large keeled terminal plates followed by a pair of narrow, long keels.
Dorsal scales: Head: 18-19 rows; body 10-11 rows with a total of 33-40 thin, strongly overlapping heptagonal to rectangular scales (19µm x 9µm), each bearing a spine (2µm - 40µm) with a secondary tip; short spiny field at the rear end
Ventral scales: ventral intermediate field with two curved, keeled terminal plates;8-9 rows of keel scales (partly spiny); oval, smooth scales in the pharynx region
Particularities: Distinct system of pharyngeal veins
Chaetonotus hoanicus is a very rare gastrotrich that has so far only been observed by Schwank. I would therefore like to describe it in more detail. The main characteristic of Ch. hoanicus is the abrupt transition from an unspined (or shortly spined) head area to a relatively long spined back:
Ch. hoanicus: Change of the dorsal spination
The scaling of the neck/head area consists of almost round, strongly overlapping scales with only an extremely short - often only perceptible - spine:
Ch. hoanicus: Change in the dorsal scales on the neck
With an abrupt transition to long spiny scales the area of the trunk scales is clearly separated. The trunk scales are relatively large, roughly pentagonal and have a large cut-out at the rear end. Spines approx. 15-20 µm long arise from its tip, which are provided with a secondary tip distally.
Ch. hoanicus: Modification of the dorsal trunk spination
In the anal region there is a short spined field. Some conspicuous, longer spines protrude into the toe cutout.
Ch. hoanicus: Change in the dorsal spination of the anal region
The exact scale shape only becomes clear when it is possible to detach scales:
Ch. hoanicus: Individual dorsal scales from the dorsal region
The particularly pronounced tuning fork-shaped structure of the pharyngeal surface, which connects the oral cavity with the glands in the pharyngeal wall (see Pharynx), is conspicuous:
Ch. hoanicus: Structure on pharyngeal surface
If one films a living specimen of Ch. hoanicus with the focus on these “veins”, a clear flow within these structures becomes visible, indicating a gland-like function:
Dorsal scales: 17-19 rows of 38-40 circular scales each with distal cutout and short and simple spines
Ventral scales: 2-3 pairs of elongated, keeled terminal plates; 5-7 rows of small, round keel scales
Oecology: very europhilous species; standing and flowing waters; moor; common
Similar species: well-defined due to stocky shape and bare toes
Particularities: Bare toes; insufficiently described
The small and common gastrotrich Chaetonotus (Ch.) multispinosus is characterized by its short-spined, roundish scales, which are cut out at the posterior end:
Ch multispinosus: ventral
The short spines that give the species its name can be seen in cross-section. The short, club-shaped pharynx is very typical:
Ch multispinosus: cross section
The ventral intermediate field is covered with a few rows of roundish, keeled scales. The posterior end is formed by two larger, short spiny plates, which are bordered by two pairs of small, also short spiny keel scales:
For the subgenus Hystricochaetonotus are the relatively large, non-overlapping scales with a long spine. This subgenus owes its name to this spiny appearance (hystix = porcupine).
The scales are in typical arrangement. Their shape is trilobate with a central keel, at the end of which arises a long spine - usually with a secondary tip:
typical three-lobed scales in Hystricochaetonotus - here
C. (H. ) persetosus
Dorsal scales: 13 rows of 13-14 three-lobed scales (6.5µm x 5µm) with simple spine (3µm - 10µm); typical scale group for Hystricochaetonotus without spines at the posterior end
Ventral scales: ventral intermediate field with two rows of 20 rectangular scales each (1.2µm - 4µm); two keeled terminal plates (length 6.4µm - 6.8µm)
Oecology: Moor
Similar species: all Hystricochaetonotus; precise diagnosis required
Particularities: Spines without secondary tip; unique scaling of the ventral intermediate field
The small gastrotrich Chaetonotus (H.) italicus, previously only known from Italy, is relatively common in the Sima bog.
Ch. italicus: dorsal
Many of the specimens found were in their post-parthenogenetic (hermaphroditic) phase and bore both a distinct, bipartite X-organ and rod-shaped spermatoids. The pronounced dumbbell-shaped pharynx is typical of the species:
Ch. italicus: t.s.
The ventral intermediate field is covered with two rows of rectangular scale plates, which become more indistinct in the neck area. Two keeled terminal plates form the end:
Ch. italicus: ventral
Although the scale spines do not have a secondary tip, the classification in the subgenus Hystricochaetonotus is clear due to the typical scale shape:
Ventral scales: 2 long, thin terminal plates with short spines; 7-8 rows of small keel scales (1µm - 3µm)
Oecology: Moor, bank moss
Particularities: Toes naked; rigidly protruding spines are very typical
The very rare gastrotrich Chaetonotus (H.) murrayi is only known from two localities (in Poland and near Berlin). The strong and quite long two-pointed spines protrude in a spear shape and give the animal a very defensive appearance.
Ch. murrayi: cross section
This is particularly noticeable when focusing on the spines on the back:
Ch. murrayi: dorsal
The approx. 20µm long spines have a lateral secondary spine near the main spine:
Ch. murrayi: focus on the spine tips
The focus on the scales shows the typical three-lobed scale shape of this subgenus. In contrast to the findings in the literature, the entire head of the animals is also covered with long spines:
Ch. murrayi: dorsal scaling of the head / neck area
The spination is also clearly pronounced at the posterior end. Unlike many members of the subgenus Hystrocochaetonotus this species lacks the unspined dorsal keel plates:
Ch. murrayi: dorsal scaling of the hind end
The ventral intermediate field bears 6 - 7 rows of small keel scales and is terminated by two spiny terminal plates.
Ch. murray: Scales of the ventral intermediate field
Dorsal scales: 7 rows on the head, 12 on the trunk, each with 14-15 three-lobed, keeled scales (3-6 µm), which do not overlap and are short and delicately spined (10 µm); two rows of longer spines at the head end. Very long spines with two secondary tips (26-50 µm) in a relatively narrow field on the anterior trunk
Ventral scales: 2 elongate-oval, keeled terminal plates (6.6µm x 4µm) with two simple spines; 4-6 rows with 40 oval keel scales each. Own observation: in pharyngeal region rectangular transverse claspers
Oecology: Moor, plant-rich small bodies of water
Similar species: C. (H. ) anomalus
is possibly synonymous; differs only by the absence of the two scale rows with longer spines on the head (which was present in my observations) and by scale claspers below the pharynx (which are also present in Ch. novenarius in my observations).)
Particularities: easily delimited by the long spines that emerge from a narrow field
Chaetonotus (H.) novenarius is immediately recognizable by its nine very long large spines.
Ch. novenarius: dorsal
The nine large spines each have two secondary tips and extend beyond the end of the animal’s toes. Ch. novenarius is the Chaetonotus with the longest spines relative to the body length (approx. 2/3 of the body length):
Ch. novenarius: dorsal large spines
Although the scales and spines clearly identify the animal as Ch. novenarius, the ventral scales of the found animal are puzzling:
Ch. novenarius: ventral
Striking are the rectangular scale plates in the pharynx area, which Schwank only shows for the very similar species Ch. anomalus. It is possible that this species is a synonym of Ch. novenarius, as (Schwank, 1990)does not rule out.
Dorsal scales: 5 rows of 15-18 transverse oval scales (5 x 10 µm) each, not touching; on head dorsally and laterally 5-7 characteristic spiny scales; small spines 6-7 µm; groups of large spines 8-22 µm.
Ventral scales: Ventral intermediate field on head 3 touching plates; otherwise elliptical plates; in gut region sometimes 3 deer of smaller elliptical plates.
Oecology: Bog waters, rare, very variable
Particularities: very variable with respect to spine size - possibly a number of subspecies
A very rare and strange ventral hardy is the nearly rotund Chaetonotus (P.) acanthocephalus with its irregular spination and large, round scales:
Ch. acanthocephalus: focus on the dorsal scale
Ventrally, the animal bears large cuticular plates on the head that change to round, short-spined scales toward the posterior end:
Ch. acanthocephalus: ventral
If the animal is macerated with glacial acetic acid, the scale shape typical for the subgenus Primochaetus becomes particularly clear:
Dorsal scales: 10 rows of 20-25 strongly overlapping, rounded triangular scales (5 µm - 8.5 µm) with strongly convex margin (margin appears double) with short spines (7 µm); 7 - 12 simple girdle spines (6.5 µm - 16.5 µm) in alternating rows; scales behind spine girdle mostly without spines
Ventral scales: 4 elongated terminal plates with keel; 8-9 rows of small keel scales
Oecology: Moor, on mud
Similar species: well demarcated by spine belt and double scale edge
Particularities: Spiked belt and double scale edge
A further representative of the subgenus Primochaetus is the small abdominal hardy Ch. (P.) acanthodes, which is notable for its distinct, spineless scales with a “double” anterior contour and a belt of long spines on the abdomen:
Ch. acanthodes: dorsal scales
Only in the lateral view one recognizes that the spiny belt covers the entire dorsal side of the animal:
Ch. acanthodes: side view
The ventral intermediate field is covered with approx. 8 rows of small, roundish keel scales. The rear end is formed by four rectangular, narrow keel plates:
Ch. acanthodes: ventral scales
In cross-section, it becomes clear that the scales are strongly curved. This results in the “double contour” in the optical section. The animals I found have one or two teeth at the entrance to the pharynx:
Dorsal scales: 7 rows of fe 20 thin, transverse-oval to obovate scales, only slightly touching each other; spines (10-39 µm) originate slightly behind scale center; secondary tip relatively far from distal end; three pairs of long lateral spines (34-46 µm) overhang toes by twice their length; between toes 3 dorsal spines slightly overhanging toes
Ventral scales: Ventral intermediate field with +11 rows of short spiny tiny scales
Particularities: brood care: decorating the eggs with detritus; var. bernensis has no secondary tips
Ch. cordiformis shows in cross section the typical structure of limnic gastrotrichs. Remarkable are the relatively few, long and strongly curved scale spines:
Ch. cordiformis: cross section
The side view shows the enormous length of the spines which are provided with a secondary tip:
Lateral view(ChCordiformis2.jpg)
Ch. cordiformis: side view
These long spines make it very difficult to take good photos of the animals, because pinning them down with the coverslip strongly disturbs the natural arrangement of the spines. Therefore, an attempt should be made to capture the appearance of the animals free-swimming:
Ch. cordiformis: free swimming specimen with focus on the scale spines
High resolution images are only possible with somewhat pressed specimens. Only here the roundish scale shape with the scale spines provided with a secondary tip becomes visible. The secondary tip sits relatively far away from the spine end of the animals.
Ch. cordiformis: spiny scales with secondary tip.
Many of the specimens found had peculiar spherical bodies inside, which I think are (permanent forms? of) parasites:
Ch. cordiformis: spherical inclusions.
The scales of the animals can be judged well only after they are detached. Here you can recognize the name-giving “heart shape” of the roundish scales with an indentation:
Ch. cordiformis: heart-shaped scales
Ch. cordiformis is one of the few gastrotrichs that performs a kind of “brood care” after egg laying. After oviposition, the mother transports detritus from the vicinity of the oviposition site in her pharynx to the egg and sticks this material to the eggshell. As a result, the eggs are completely covered by detritus and protected by it from enemies:
Ch. cordiformis: brood care - start movie by clicking.
Dorsal scales: 5 Reihen mit je 20 einfachen, dicken und kurzen Stacheln; lateral 12 Großstacheln: 2 Paar am Hals, 1 Paar in der Mitte, 3 Paar am Hinterende
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Schimkewitsch,W. 1886. Uber eine neue Spezies Ichthydium. Nachr. Gesell. Freunde Naturw. Moskau 50: 148-150.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus crinitus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: crinitus
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: unzureichend beschrieben und so nicht wieder zu erkennen!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Sudzuki,M. 1971. The Gastrotricha of Japan which live in the capillary water of the interstitial system: II. Bull. Biogeogr. Soc. Japan 27: 37-41.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus dentatus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: dentatus Tretyakova, 1992
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Chaetonotus elegantulus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: elegantulus Remane, 1936
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: sp. inquirenda; jegliche Abbildung und Beschreibung fehlt!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Remane,A. 1936. Gastrotricha, In: H.G. Bronns, ed. Klassen Ordnungen des Tierreichs, Band 4, Abteilung II, Buch I, Teil 2, Lieferungen 1-2. Akademie Verlagsgesellschaft, Berlin.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus gracilis
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: gracilis Gosse, 1864
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: taxon inquirendum; keine Angaben über Schuppen und Stacheln, daher nicht wieder erkennbar
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Gosse,P.H. 1864. The natural history of the hairy-backed animalcules (Chaetonotidae). The Intellect. Observer 5: 387-406.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus hermaphroditus
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: hermaphroditus Remane, 1936
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Particularities: taxon inquirenda; jegliche Abbildung und Beschreibung fehlt!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Remane,A. 1936. Gastrotricha, In: H.G. Bronns, ed. Klassen Ordnungen des Tierreichs, Band 4, Abteilung II, Buch I, Teil 2, Lieferungen 1-2. Akademie Verlagsgesellschaft, Berlin.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus lancearis
Taxonomy
Order: CHAETONOTIDA Remane, 1925
Suborder: PAUCITUBULATINA d'Hondt, 1971
Family: CHAETONOTIDAE Gosse, 1864
Subfamily: CHAETONOTINAE Kisielewski, 1991
Genus: Chaetonotus Ehrenberg, 1830
Species: lancearis Tretyakova, 1992
Length ( flaschenförmig ):
Width:
Width of the head ( fünflappig ):
µm
Length of the furca:
Adhessive tubes:
Pharyx ( zylindrisch ):
Diameter of the mouth ( rund ): unknown
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Dorsal scales: dorsolateral 3 Bündel mit leicht gebogenen, abspreizbaren Stacheln mit je zwei Nebenspitzen; (Hals: 3 x 130-140µm, 2 x 35-38µm; Mitte: 3 x 80-90µm; Hinten 2 x 60-70µm)
Dorsal scales: Dorsolatereal 9 Paar s-förmig geschweifte Stacheln mit 2 Nebenspritzen in 3 Bündeln (Hals: 3 x - erste zwei 180-182µm, dritte 40µm; Mitte: 4 x 130-132µm; Hinten: 2 x 60-72µm)
Dorsal scales: Hinten: 2 stumpfe Lappen mit kurzem Stachel (17-20 µm) mit Doppel- und Nebenspitze; auf einer Seite 12, auf anderer Seite nur 3 z.T. riesige Stacheln mit Doppel- und Nebenspitze; 3 dünne Stacheln am Hinterende (50µm)
Ventral scales: 4-5 Paar Zilienbüschel, sonst nackt
Oecology: über Faulschlamm, Lemnatümpel
Similar species: durch asymmetrische Stacheln einzigartig
Dorsal scales: Auf den Hinterlappen ein Paar langer, auf Schuppenrudimenten sitzende Stacheln (63-68 µm; 1/4 Körperlänge); 2x7 Stachelbündel mit proximaler Nebenspitze; Kopf 2x mit 1 bzw 3 Stacheln; Rumpf 5x mit 4-7 Stacheln; Stachelanzahl und -länge (60.76µm) variiert
Ventral scales: 5 Paar Wimpernbüschel
Oecology: semiplanktisch über Schlamm
Similar species: evtl. synonym mit
D. (D. ) ornatus
; Unterschied Stacheln
Particularities: sehr viele gleichmäßiger verteilte Stacheln
Dorsal scales: Kopf zwei Ringe mit je 6 Stacheln mit einer Nebenspitze (10-15 µm); Körper 4 Paare Stachelgruppen mit je 5 Stacheln mit zwei Nebenspitzen (69-115 µm); Hinten zwei Paare Stacheln mit zwei Nebenspitzen (18µm); zwei Paar einfache Stacheln
Dorsal scales: neun Paare Stachel-Gruppen; Stachelzahl von vorne: 1 - 1 - 2 - 5 - 5 - 3 - 1 - 1 (58-75 µm); erste beiden Gruppen einfach, sonst zwei Nebenspitzen; auf Schuppenrudimenten; Schlägerartige unpaarige Struktur am Hinterende (59x17 µm), kann fehlen (vielleicht epizooisch)
Oecology: zwischen Wurzeln; Brasilien
Similar species: D. (D. ) ornatus
hat weniger Stachelgruppen und keinen “Schläger”
Dorsal scales: terminal 2 dick Stacheln (78-85 µm), distal mit Doppelspitze, an der proximalen Nebenspituze stark abgewinkelt; 8 Paare Stachelbündel: Kopf 1x; Hals 2x; 6 Paar Rumpf 3-5 x; längste stacheln 84-86 µm; am Kopf 3 weiter Stacheln, mittlere ohne Nebenspitze
Dorsal scales: No scales; 5 pairs of non-crossing elastic spines projecting above the body; two spines each starting ventrally
Ventral scales: no scales; 3 pairs of long jump spines (100 - 240 mm); posterior ones overhang the body by one body length and cross each other
Oecology: over mud, formerly frequent, meanwhile rare
Similar species: H. festinans
: lower number of jump spines
Particularities: Setae stand on clear papillae
Haltidytes crassus was a relatively common species at the time of its first description. Since then - probably due to the influence of industrial agriculture - this pretty gastrotrich has become very rare. The main characteristic of H. crassus are the five dorsal and three ventral “jumping spines”, with which the animal catapults itself out of the danger zone when threatened.
Dorsal; ds: dorsal spines (5 pairs).
The long, caudal and hair-shaped last ventral pair of spines, which far overhang the body and cross each other in resting position, is conspicuous.
Three ciliary rings on the head are used for swimming locomotion, while the ventral ciliary bands typical of gastrotrichs are used for gliding over the substrate.
Ventral; cr: 3 ciliary rings for locomotion; js: jump spines; cs: long caudal spines; se: seta on papillae.
The posterior palps - as in all members of the genus Hatidytes - are on distinct papillae.
Median; br: “brain”; ph: pharynx; oo: mature ovum; in: “intestine”; mf: muscle fibers that move the jumping spines.
The internal anatomy has some peculiarities. For example, distinct muscle fibers can be seen that couple to the attachment sites of the jumping spines. Adhesive glands and the associated adhesive tubes are not present in this genus.
The typical movement and behavior of the animals can be seen in the following YouTube video:
Dorsal scales: no scales; dorsolaterally 3 groups each of 3 + 2 + 2 jump spines, which protrude even in resting position and slightly overhang the body
Ventral scales: no scales; a pair of long jumping spines (140-144 µm), surpassing the body by 3/4 of its length and crossing each other
Oecology: About digested sludge
Similar species: H. crassus
: more ventral spines, less lateral spines
Heterolepidoderma macrops is one of the most common and widespread Heterolepidoderma species and was certainly often confused in the past with
H. ocellatum
from which it differs - apart from some details that are difficult to see - mainly by the larger number of scales:
Dorsal view; oc: pseudocells; ss: sensory scales.
In most animals, the pseudocells resembling eyes are very conspicuous, but they have no sensory function. The posterior sensory hairs are located on small, triangular “sensory scales”.
Dorsal with focus on the dorsal terminal scales (dts).
The toes of the animals are unscaled. The scale ends with a pair of relatively large keel scales at the base of the toes.
*Cross section
In the section you can see that the small scales - in contrast to
H. ocellatum
- do not bear a distal process. The pharynx is slightly swollen on both sides.
Central view with terminal plates (tp)
Ventrally, the animal bears only a pair of small, oval terminal plates. The remainder of the ventral interfield is unscaled.
The adhesive tubes are thickened terminally.
Staining of nuclei with methyl green; B C D: Nuclei of the brain; A: overview.
If H. macrops is stained with a selective nuclear dye like methyl green, only the nuclei of the animals are stained. This gives a good overview of the structure of the brain. Clearly visible in subfigure B is the nucleus-free area dorsal to the pharynx, which is referred to by (Zelinka, 1889)and (Remane, 1936)as the “dorsal commissure”, i.e. a nucleus-free connection of the two cerebral hemispheres made of nerve fibers, and which is also confirmed in more recent studies (Schmidt-Rhaesa and Holger Rothe, 2014). Lowering the focus to the midplane of the pharynx (subimage C), one recognizes the pharynx consisting of a few cells surrounded on both sides by brain cells. Ventrally, no connection of the two cerebral hemispheres can be seen below the pharynx. Thus, the brain does not completely enclose the pharynx and delimits the animals to the Cycloneuralia. The already unstained conspicuous pseudocells prove to be stainable with methyl green - a strong indication that these structures are the prominent nuclei of the sensory neurons of the tactile hairs.
Scale preparation with eosin.
The scale shape of the small dorsal keel scales of H. macrops cannot be judged exactly in the scale compound on the living animal. A more precise [scale preparation](/technique/scale preparation) clearly shows the oval shape of the scales, which - in contrast to
H. ocellatum
- do not wear a distal tip.
After a phase with parthenogenetic reproduction, the animals enter the so-called “postpathenogenetic phase”. In this stage of life, the animals reform into hermaphrodites and simultaneously develop eggs and sperm produced in bundles ventrally to the side of the intestine - they reform into true hermaphrodites.
In H. macrops the spermatoids are spindle-shaped. At the same time, a bipartite, so-called X-organ with unknown function is formed. Here the X-organ consists of two spherical, secretion-filled cysts connected by a bar (or connecting duct).
Heterolepidoderma majus is a very widespread, slender and elegant belly-hardy.
H. majus: dorsal scaling ( Ole Riemann)
One of the animals observed was in its post-parthenogenetic (hermaphroditic) phase and showed a sperm bundle in addition to the conspicuous bipartite X-organ at the posterior end:
H. majus: x-organ (xO) and sperm bundle (Sp)( Ole Riemann)
The scaling at the base of the toes is typical for the species. The sensory hairs arise from special scales:
H. majus: Scaling of the base of the toes ( Ole Riemann)
The ventral intermediate field is occupied by small, elongate keel scales and bears two terminal keel plates:
H. majus: Scaling of the ventral interfield
Another feature that makes the species distinctive is the demarcated “stomach ring” on the foregut:
Similar species: Ichthydium monolobum was included in this species
Particularities: Very characteristic furca
The species I. (F.) forficula was united some time ago with the somewhat smaller species I. monolobum into a single species. A main characteristic of this species are the long, curved and thin glue tubes, which give the animals a “tweezer-like” appearance:
I. forficula: dorsal with pincer-like adhesive tubes
On the ventral side the animal carries two distinct terminal plates.
I. forficula: ventrale Terminalplatten
The lash bands are broken up into individual clumps of extremely long lashes.
I. forficula: Ciliate tuft
In the side view, you can see the enormous length of the locomotive eyelashes:
Dorsal scales: only triangular basal scales of terminal setolae
Ventral scales: naked except for two rectangular, unkeeled terminal plates; rows of cilia on trunk resolved into 4 pairs of ciliate tufts; bands of cilia united on head
Oecology: In sphagnum / bog
Similar species: I. (F. ) skandicum
: Additional dorsal scales at posterior end
Particularities: Very long dorsally visible cilia (27µm); very rare, only a few specimens in Italy and Sweden so far.
Ichthydium (Furficulichthys) tanytrichum is a very rare belly-harper, which since its first description in Italy in 1982 (Balsamo, 1982)has only been reported from Sweden - as a single specimen (Kånneby et al., 2009). The here shown finding from the Austrian Lauchseemoor is the third worldwide known occurrence of this species.
With a size of about 130µm the animals of the found population are about the same size as the animals found in Sweden. The Italian animals of the first description were significantly smaller (ca. 74µm). Already Kånneby expressed the suspicion that the first description was based on juvenile animals, because the adult animal found by him was clearly larger with 127µm. This would also explain the somewhat different proportions to the first description.
Median cut
The lack of a continuous scaling and the flexible cuticle are characteristics of the genus Ichthydium. The typical pincer-shaped furca of the animals places I. tanytrichum clearly in the subgenus Furficulichthys, within which the species can be clearly defined by body shape and lack of scaling.
Dorsal: “sensory scales” from which the tactile hairs arise.
Other species characteristics are the triangular “sensory scales” from which the posterior tactile hairs arise. The anterior pair of tactile hairs sits on small papillae. Further scales are not present on the dorsal side.
Ventral: In the ventral region, the cilium bands are resolved into single tufts.
On the ventral side the long cilia are noticeable, with which the animal moves. With a length of about 25µm they give this species its name (“tanytrichum” = “long haired”). Typical for this species is, that these locomotive cilia are not organized in two continuous cilium bands - as otherwise common in gastrotrichs - but stand in single cilium tufts. Terminal are faintly recognizable the two large rectangular terminal plates.
One of the animals found had reached its postpathenogenetic (hermaphrodite) phase and was already carrying an X organ and sperm bundle:
L. minor chaetifer: above dorsally with smooth scales typical of Lepidodermella;
middle in cross section with X-organ and sperm bundle;
below ventral scaling with distinct terminal plates ( Ole Riemann).
In Sima Marsh the somewhat divergent subspecies Lepidodermella minor chaetifer was found, differing from the species L. minor minor by a pair of spines at the base of the toes:
L. minor chaetifer: the pair of spines at the base of the toes is characteristic of the subspecies chaetifer
Another striking feature of this species is the single tooth projecting asymmetrically from the oral cavity:
Dorsal scales: 7-9 rows of 25-30 very differently shaped, spikeless, strongly overlapping scales; head and trunk scales elongated oval with triangular / broadened front end (12 x 10 µm); neck scales transversely oval, short (5 x 9 µm); hypostomium: two cusps
Ventral scales: ventral intermediate field with more than 20 transverse claspers below pharynx and 3-5 rows of quadrangular rounded, overlapping scales; 2 large terminal plates and some small, round scales; ciliary bands anteriorly mostly united
Oecology: Frequent, found everywhere
Particularities: well delimited by rectangular scale plates and large terminal plates; very variable and well studied species
Lepidodermella squamata is one of the best-studied gastrotrichs, as it is kept in culture in many laboratories around the world. However, the “cultivated forms” often differ significantly from the “wild forms”, as the animals, with their enormous generation rates of around 50 per year, can react quickly to the adaptive pressure of the habitat. L. squamata is therefore considered a very variable species. The photos shown here are of wild forms.
Dorsal scales
Ventral scales; large terminal plates; rectangular braces under neck
L. squamata is one of the many species of gastrotrichs in which the glandular ducts on the surface of the pharynx are clearly visible. These ducts connect individual gland cells in the pharyngeal wall and channel the secretions of the glands into a common excretory duct that pours into the animal’s oral cavity:
Width of the head ( five lobes, broad ): 38 µm - 40 µm
µm
Length of the furca:
Adhessive tubes:
Pharyx ( cylindrical ): 35 µm
Diameter of the mouth ( around ): unknown
Dorsal scales: posterior setolae on small keel scales; terminal spine 45µm; spines in 5 groups of one short ventral spine (10-15 µm) and one longer lateral spine (ca. 40 µm); anterior and last group unpaired, i.e. 8 spines per side
Ventral scales: naked; only one pair of eyelashes; eyelash tufts on head typical
Oecology: plant-rich pools; over mud; rare
Setopus bisetosus (Thomson, 1891) - a formerly common species - has become very rare due to the progress of industrial agriculture and the accompanying drainage. Therefore, it always pleases me to find a specimen of this species.
free swimming copy
Unfortunately, I could not completely still the animal, so the photo quality was not optimal.
The semi-planktic Setopus species usually do not have continuous ciliary bands and move swimming with an (almost completely closed) ciliary ring on the head. In addition, some ciliary tufts are present ventrally on the head. On the body itself, except for a few isolated cilia, the ciliary bands are reduced to a pair of ciliary tufts on the abdomen.
In median section, two pairs of keeled terminal plates can be seen at the posterior end of the animal. These terminal plates are not mentioned in the literature.
Laterally five pairs of spines are visible.
Median section with terminal plates (tp) and head shield (kep); 1..5: lateral spines; cilia ring (cr).
Typical is the broad and strong head shield (kephalion) of the animals.
Dorsal scales: Terminal spines with secondary apices on triangular scale rudiments; unequal in length (27-30 µm; 43-55 µm); numerous dorsolateral spines with secondary apices on scale rudiments; 21 per side.
Oecology: rare; bog lake
Similar species: S. chatticus
: no scales and fewer shorter spines
Particularities: scaled ventral interciliate field
Setopus tongiorgii has been found so far only in Italy and some locations in Germany and is - like all Setopus species - relatively rare.
Overview with focus on the ring of eyelashes on the head ( Dr. Stephan Krall).
Transverse section; Rs: trunk spines; Cs: caudal spines ( Dr. Ole Riemann).
In the cross section the lateral spines with their secondary spike are well visible. Terminal are two caudal spines of unequal length, which also carry a secondary tip.
Scaling of the ventral intermediate field ( Dr. Ole Riemann).
Ventrally, the small, roundish scales and the keeled terminal plates of the ventral interciliar field can be seen. So far only in S. tongiorgii scales were found in this area. This feature is therefore of special importance for the diagnosis of the species.
With belly-harriers the species classification is traditionally done by the shape of the scales. Therefore, species identification is often only possible if one can accurately assess the scale shape of the animals. Unfortunately, the scales of gastrotrichs are often difficult to see. Even with DIC - although here the weak birefringence of the scales further enhances the contrast - live observation is not sufficient for species diagnosis in many species. Therefore I would like to present here a possibility for the preparation of the scales and thus for the (mostly) safe species diagnosis. As an example I have chosen
Lepidochaetus zelinkai
. Due to the distinctive appearance of this gastrotrich, a scale analysis for species identification is actually not necessary, but some scale details are controversially discussed in the literature, so I wanted to make up my own mind.
L. zelinkai
is a medium sized gastrotrich with a distinctive spiny coat:
To prepare the scales free, the animal must be macerated. To do this, fix the animal - preferably with a simple Vaseline copressorium - with the coverslip and draw a drop of glacial acetic acid under the coverslip. I stained the glacial acetic acid in advance with very little eosin - this stains the scales at the same time as maceration. The glacial acetic acid dissolves all soft parts of the animal within a few minutes, while the scales of the gastrotrichs are acid-resistant and therefore remain intact. Depending on the scale structure, the scales remain as a cohesive shell or separate from each other. In
L. zelinkai
, the scales are apparently not interlocked, so the scales disperse:
This type of preparation is quite simple and allows the scale shapes to be accurately assessed. For example, in the above images, the ridges in the scale surface that extend parallel to the leading edge are noticeable. In the literature, these “double edges” of the scales are usually considered to be an optical artifact produced by the double intersection of the focal plane through the curved scales. The above scale analysis clearly proves that these conspicuous ridges really exist and represent a thickening of the scales.
Microaquarium
By microaquaria I mean slide-sized preparations that can be observed directly with a microscope. The optical quality of the microaquaria (especially the thickness of the slides) should be good enough to use any possible magnification. In addition, the life span of the organisms inside should be at least a few weeks to be able to study the behavior or development of the objects. Last but not least, the method must be easy and fast to use - a cumbersome method that requires a lot of equipment is not practical in my eyes in the amateur field. A few years ago I came across a preparation method on the Internet, which I was able to develop into a long-term microaquarium. Such a microaquarium is constructed as follows:
Construction of a microaquarium
The life of a “normal” temporary wet preparation can be increased by effectively preventing the water from evaporating. This can be accomplished by sealing the cover glass with a Vaseline rim to make it watertight. Vaseline has a number of ideal properties for this purpose:
it is absolutely waterproof
Vaseline is permeable to air (known from Vaseline-based moisturizers)
Vaseline is easy to apply due to its consistency
it can be found inexpensively in every drugstore
To apply a Vaseline edge to the coverslip, apply a thin (!) film of Vaseline to the heel of your hand and wipe a thin ridge off the skin with each coverslip edge. The coverslip can then be placed directly on the slide (with specimen) with one edge. From this edge, lower the coverslip and slowly press the coverslip to the desired thickness using a wooden stick. Excess water is thus pressed out from under the coverslip and can then be easily removed.
To create a long-lasting microaquarium, it is necessary to include a relatively large amount of water in the preparation. If this water buffer is too small, the organisms would die after a few days. Therefore, a microscope slide with a hollow section is used for a microaquarium. To increase the area that can be used for observation, I use a coverslip that is as large as possible (e.g. 24mm x 32 mm or 24mm x 60mm). This maximizes the area outside the hollow grounded section - the area with the optimal slice thickness.
A microaquarium created this way has many advantages:
Lifespan of organisms up to 6 weeks
great mechanical stability:
cleaning is easy due to the fixed cover glass
vertical storage in normal preparation boxes (transparent cover!)
absolutely stable conditions also for long-term video recordings
Use of immersion oil is unproblematic: cover does not slip, oil does not penetrate under the cover glass
additional effort for preparation only a few hand movements
But the main advantage in my eyes is that the organisms behave “naturally” after an acclimatization period of about one day in the micro aquaria. The typical great restlessness known from the “normal” temporary wet preparations has subsided and the enclosed animals go about their “daily business”. Most ciliates calmly graze their food or even stand in the same spot for hours, swirling their food in a relaxed manner. Gastrotrichs calm down and emerge from their hiding places after some time and graze - optimally observable - the underside of the cover glass. To demonstrate these relaxed conditions in a microaquarium, I would like to show the following - almost unedited film - taken in several microaquariums a few days after their creation:
Please click on the picture to start the movie!
For me this kind of preparation is meanwhile the most satisfying way to observe the wonder world in the pond. Of course it is not possible to show the last anatomical details of the organisms. But you get the possibility to observe the behavior or the development of the organisms over a long period of time and without stress.
Coverslip Compressorum
To observe living gastrotrichs in detail microscopically, it is necessary,
to minimize the slice thickness, as this is the only way to ensure optimal optical conditions for objectives with high magnifications
to keep the animals immobile
to minimize the drying of the specimens to the extent that a comfortable observation time can be achieved
prevent the penetration of immersion oil into the specimen
For this purpose I have developed the coverslip compressorum, which allows to achieve these conditions very easily and quickly.
First, as always, it is necessary to isolate the observation object with a thin pipette under the stereo microscope and apply it to a cleaned slide with as small a drop of water as possible. Care should be taken to ensure that the water droplet is small enough so that the layer of water under the coverslip to be applied does not reach the edges of the coverslip at the desired thickness. Some experience is required for this…
Coverglass compressorium: vaseline bars in yellow.
Some vaseline is rubbed on the heel of a hand. With the upper and lower edge of a rectangular cover slip (I usually use the size 18x18mm) one strips a thin bar of vaseline from the ball of the hand. The vaseline serves as a spacer around the edge of the cover glass. The coverslip is placed on the slide with one edge at an angle - the Vaseline holds the coverslip in this position. Using a small wooden stick, the protruding edge of the coverslip is carefully pressed towards the slide until it reaches the water drop and also rests with the upper side on the slide. The water drop now fills a small section of the coverslip with a relatively large layer thickness.
In this state, the gastrotrich trapped in the water droplet is still free to move and can be conveniently observed with weaker objectives. The small area of the water drop makes it easy to find the object.
Under microscope control, the layer thickness of the water can now be carefully reduced by pressing the coverslip edges just enough so that the gastrotrich is no longer movable and is gently pressed through the coverslip onto the slide. The two Vaseline edges provide sufficient resistance to manual pressure to reduce the coating thickness in a very controlled manner.
The two opposing Vaseline edges ensure that
drying of the preparation is greatly delayed, since evaporation of the water only occurs on the exposed coverslip edges without Vaseline and is therefore reduced.
the adjusted layer thickness remains almost unchanged when the preparation dries out, since the water drop evaporates only at the edges and only its diameter changes.
penetration of immersion oil is impossible at the sealed coverglass edges, and there is usually no contact with the sample water at the exposed ones.
a “pull-through” of chemicals for dyeing etc. is possible without problems because of the open cover glass edges on the opposite side.
a later manual reduction of the layer thickness until the object is flattened (e.g. for scale analysis is possible at any time in a controlled manner.
With a little experience, the creation of a cover glass compressorum is easy and possible within a few seconds.
I have been using this preparation method for high-resolution observations for years and would not want to do without it!
Double Coverslip
Gastrotrichs are animals in which the upper and lower sides differ greatly. It is often necessary to observe both the ventral and dorsal sides closely. When observing an animal with a coverglass compressorium, usually only one side can be observed accurately. The other side is only accessible when focusing through the animal. However, this results in great optical quality losses - especially with high-resolution lenses. One can then only try to rotate the animal by moving the cover glass. Unfortunately, this method only has a chance of success with round objects and usually ends lethally with gastrotrichs… :cry:
diagonal double cover glass
vaseline bridge in yellow
To allow bilateral observation, one can enclose the object between two coverslips and turn the entire coverslip stack on the slide if needed. To do this, use two different sized coverslips. The larger coverslip (e.g., 20x20 mm) is pinned to a slide with a small drop of distilled water. The coverslip should be oriented diagonally so that it can be easily grasped at the opposite corners if necessary. A second, smaller coverslip (18x18 mm) is fitted with a vaseline ridge on two edges (as described in the article coverslip compressor and pressed onto the lower coverslip with the sample in a controlled manner, thus holding the object between the two coverslips.
The object can now be observed with the usual optical quality on the top side. Once observation from this side is complete, a drop of immersion oil is placed on the coverslip (if not already done) and the coverslip stack is detached from the slide at the protruding corners. The stack is rotated and placed with the oiled side on the slide. Now the observation can be made from the now upper, former bottom side. The use of different coverslip sizes ensures that only one coverslip is touched when the coverslip stack is rotated and that the coverslips do not shift against each other.
One of the main problems in the preparation of gastrotriches is to track down the animals in a water sample and pipettate on an object carrier.Since gastrotriches are transparent, they are almost invisible in a stereo microscope with traditional incident or transmitted light illumination.The animals only become visible when the field lighting is dark.
In the professional area, dark field lighting is realized by cold light conductor. LED lights with swan neck offer an alternative option that illuminate the sample on the side.This lateral lighting only gets the light spread on the objects into the lens of the microscope.The background remains pure black.Since the lamps are not attached to the sample, the illuminance and the shadow throw changes depending on the sample position.Often the lamps also represent a mechanical obstacle when shifting the sample vessel.
This type of lighting is usually not available in stereo microscopes and must be retrofitted.
In order to minimize these disadvantages, I have developed a side dark field lighting for this purpose, which can be easily realized with simple means and a 3D printer and can be adapted to any stereo microscope.
Holder for LED strips
Rotatable object guide
To align slides according to the DIC gradient or the image edges of the camera, it is necessary to rotate the slides on the stage. If you - like me - do not have a rotation stage at your disposal, you have to come up with something…
I have constructed a rotatable object guide, which can easily be manufactured with a simple 3D printer. The following properties were taken into account:
low height - thus collisions with the objectives are excluded
mounted to the stage, so that the specimens can still be moved
specimen rests directly on the stage - no problem with focusing the condenser / Köhler is possible
easy insertion and removal of the slide
different dimensions of the slides possible due to spring holder
insertion of the commercially available plastic Petri dishes with 7 cm diameter possible; additional circular recess for the use of the lids of the Petri dishes (whose height is somewhat smaller, so that no collision with the 10 objective can occur)
The object guide consists of two separate parts:
a rotating frame which is fixed to the stage with the usual fixing screws
a turntable which is placed in the frame and can be rotated in it. To facilitate the rotation, the turntable rests on the frame only at six points. The turntable is equipped with six handles so that it can be easily operated from all sides
Fig. 1: Individual parts of the specimen guide; on the left, frame that is mounted to the stage; on the right, the turntable that holds the slide or Petri dish.
The two parts are inserted into each other. The slide sits firmly in the recess and can be rotated 360 degrees and oriented as desired. The round recesses are provided for Petri dishes, if necessary.
Fig. 2: assembled object guide
At the microscope the whole thing looks like this:
Fig. 3: Rotatable object guide on microscope
This design has proven itself in practice. Of course, the rotary stage cannot be centered when rotating, the position of the specimen must be readjusted by the stage. The slides fit tightly enough so that there is no “slippage” at all when moving them. However, some slide marks seem to have a slightly smaller depth, so one wishes they would sit a little tighter on those. If you want to use such slides, you have to adjust the retaining springs a bit….
The corresponding print files - including the CAD files in OpenSCad format can be downloaded from Thingiverse. The design is adapted to the stage of my Ortholux II. For other microscopes, the position or diameter of the holding screws may have to be adjusted.
I would not like to miss this practical object guide anymore. Only by rotating the object to the optimal angle is it possible to achieve, for example, optimal contrasting with DIK. Also the alignment at the photo edges saves the constant turning of the camera.
Flash Light
Dhe advantages and disadvantages of a microflash setup on the microscope have already been discussed in detail in several articles. For me as a “Enthusiast of the Pond”, the decisive factor was that a microflash
allows to observe with minimal light intensity (of the pilot light). Unfortunately, there are many organisms that do not tolerate well the often high light levels required for DIC and panic to avoid the light or even do not survive the illumination.
ensures a constant white balance of the photos independent of the light intensity of the pilot light.
allows short exposure times independent of the pilot light. If one wants to realize short exposure times with the “normal” LED illumination, one typically has to set the lamp to maximum light intensity before releasing the shutter, but this is uncomfortable for observation. This is the only way to then achieve exposure times shorter than 1/500 s, which is necessary to “freeze” the motion blur.
Motion blur minimized with typical exposition times of less than 1/2000 at ISO 100. With LED lighting, I typically achieve only 1/200 s at 100 DIK at ISO 400.
That’s why I constructed flash equipment for my Ortholux II with my 3D printer!
With a microflash, in principle, you have the fairly simple option of mirroring the flash to the side under the condenser. However, such a setup is rather provisional, since during “normal operation” the flash and beam splitter under the object stage interfere and limit the ergonomic operation. For a permanently attached microflash on a microscope with a separate lamp house, the “flash cubes” have proven themselves. They are inserted between the lamp house and the stand and here reflect the flash into the “normal” beam path. This also allows flash illumination under “Köhler condition”, since the existing light field diaphragm is also used for the flash. So it had to become a similar flash cube solution, where the necessary mechanical components should be created with a 3D printer. My solution looks like this:
The central component is the beam splitter holder (4), which would be designed to swivel. This makes it possible to remove the beam splitter from the beam path and also to operate the illumination beam path without interference from the flash. To ensure a secure hold of the swung-out beam splitter, the swivel handle (10) was also attached to the swivel axis with a spring, so that it presses against the side wall and locks into the recess in the upper position. To swivel in the beam splitter, the handle must be pulled outward to allow free movement. On the lamp and stand side, the cube walls (6 and 7) were designed according to the dimensions for the lamp bayonet on the Ortholux II. This allows the flash cube to be firmly mounted between the lamp and stand and does not further interfere with the operation of the microscope. The light aperture (2) on the lid of the flash cube has been adapted to the dimensions of the collector lens so that it can simply be inserted directly. Therefore, when transporting, the collector lens should be removed, as it is held in place only by gravity. Due to this construction, the flash sits directly on the collector lens and the distance to the flash tube is almost optimal. The Fresnel plate of the flash cannot cause any interference in the image, as it sits very close to the collector lens and is therefore not in focus. A holder (1) for the flash is screwed onto the top, which is of course adapted to the dimensions of the flash unit used.
Assembled, the whole thing looks like this:
overall view
The outer dimensions (without flash holder) of the flash cube are 90mm * 100mm * 105mm - so it is actually a “flash cuboid”. These dimensions are due to the dimensions of the purchased parts (beam splitter, collector lens, flash) and the specifications of the lamp bayonet. In reality at the microscope, the microflash device looks like this:
Flash on Ortholux II
As a flash I use a Godox TT685II for Canon cameras with wireless remote control, which works without problems with my EOS700D. The flash has a guide number of 60 and thus enough power to still expose correctly with 1/8 of the maximum power at the 100 DIC. I operate the flash in TTL mode, which easily adjusts the flash power to the subject / lens. Only an exposure compensation of -1.7 EV had to be set. This exposure compensation is independent of the lens used, but sometimes needs to be adjusted to the subject - just like with normal photos. To operate the flash with a power supply, I replaced the batteries with battery dummies, to which a 6V power supply is connected. For those who want to try a replica, I have put together the corresponding OpenSCAD files here for download. An adaptation to the geometry of other beam splitters, collector lenses and flash units is possible without any problems. More difficult is the adaptation to lamp bayonet microscope manufacturers. This is possible in principle, but may require changes to the beam delivery, as the position of the light axis may then change. If you have problems, just ask - everything is possible somehow…
I am very satisfied with this microflash solution and would not want to miss it anymore. Unfortunately, this eliminates another excuse for my photo quality - now I guess I have to learn how to take good photos after all…
Webdesign
The plan to create a website that offers assistance in the identification of gastrotriches and systematically summarizes my own observations and findings is already several years old. When I started working on this, I - like probably most amateurs - gained my experience with WordPress.
WordPress is a wonderful content management system, that is very easy to use - as long as you follow the guidelines and procedures that WP provides. Unfortunately, I had different ideas for my website than WP’s developers and very soon I found that I was spending more work on bending WP and customizing the design than I was on incorporating content. Each new species web page had to be incorporated into appropriate navigation menus, each change to the structure of the site entailed extensive changes to WP listings. Additionally, I found it difficult to maintain a consistent web design for the individual pages, as the design was a direct part of the content of the web pages. If a decision was made at a later date to display the individual art pages differently, all existing pages had to be adjusted. At some point I had to admit to myself that WP was not suitable for my way of working and I would either have to change my workflow or give up the advantages of WP.
By chance I came across the website generator “HUGO”, which allows to generate static websites locally, which are then copied to a simple webserver only at the “push of a button” - no complicated database system, no design defaults, no proprietary plugins like with WP. And the best thing about it: HUGO tries very successfully to strictly separate content and design!
In HUGO you choose a “theme” as initial design and customize it once to your needs. The individual pages are then created in a simple text editor in “Markdown” format and generated according to my design specifications and integrated into the website. It is possible to maintain data fields in the text files, the contents of which are then included in the presentation of the website. For example, it is possible to fill out a simple, freely definable “data form” for each species of gastrotrichs, in which general size information and descriptions of the species are entered. This information is then used by HUGO to generate the corresponding web page with a standard design, e.g. for a “species profile”. Data and design are completely separated from each other. All web pages use a consistent design, which can be changed at any time for all web pages at the same time - without having to adjust all individual pages! Add to that the advantages of using static web pages, which offer unmatched responsiveness at minimal cost with full control over data privacy.
For me, using HUGO has greatly facilitated the creation and maintenance of my website and I would like to extend my sincere thanks to the many volunteer developers who have created and continually maintain HUGO :heart: !
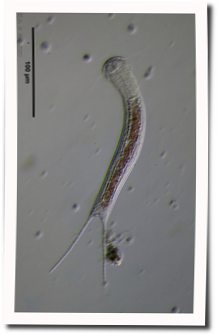
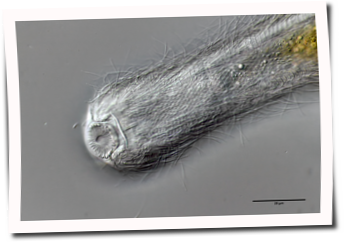




 #
#
 Dr. Stephan Krall
Dr. Stephan Krall Dr. Martin Kreutz
Dr. Martin Kreutz
 Dr. Martin Kreutz
Dr. Martin Kreutz
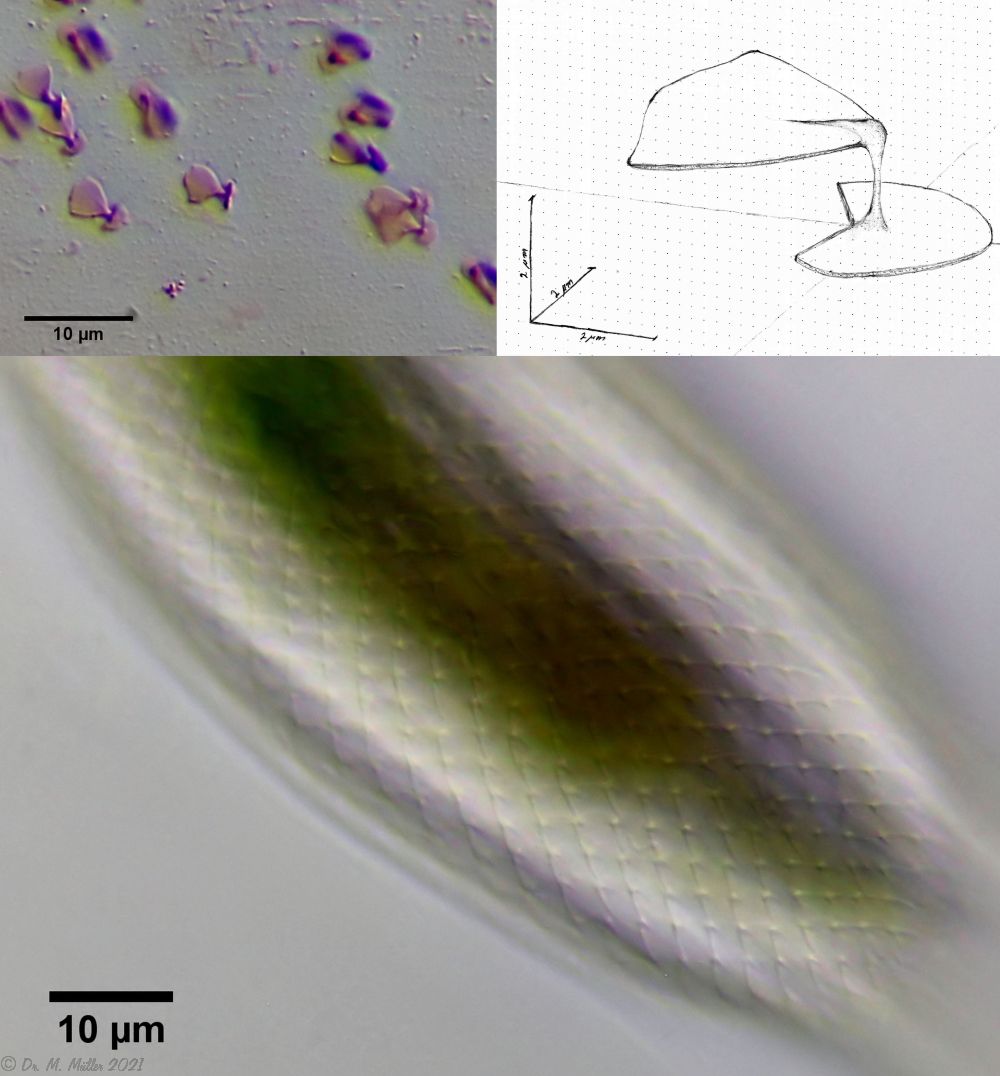 typical scales of an Aspidiophorus, here A. squamulosus.
typical scales of an Aspidiophorus, here A. squamulosus.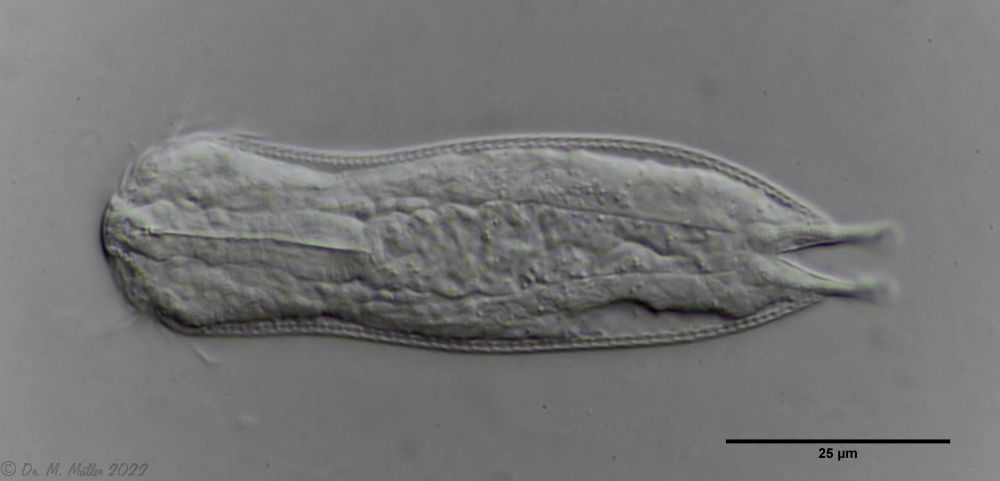
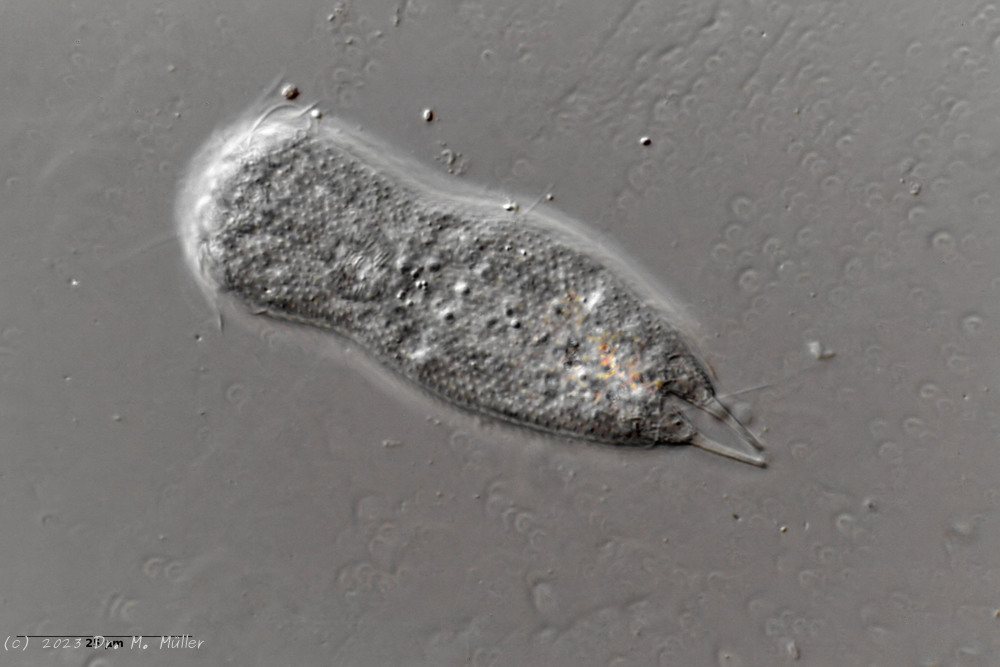
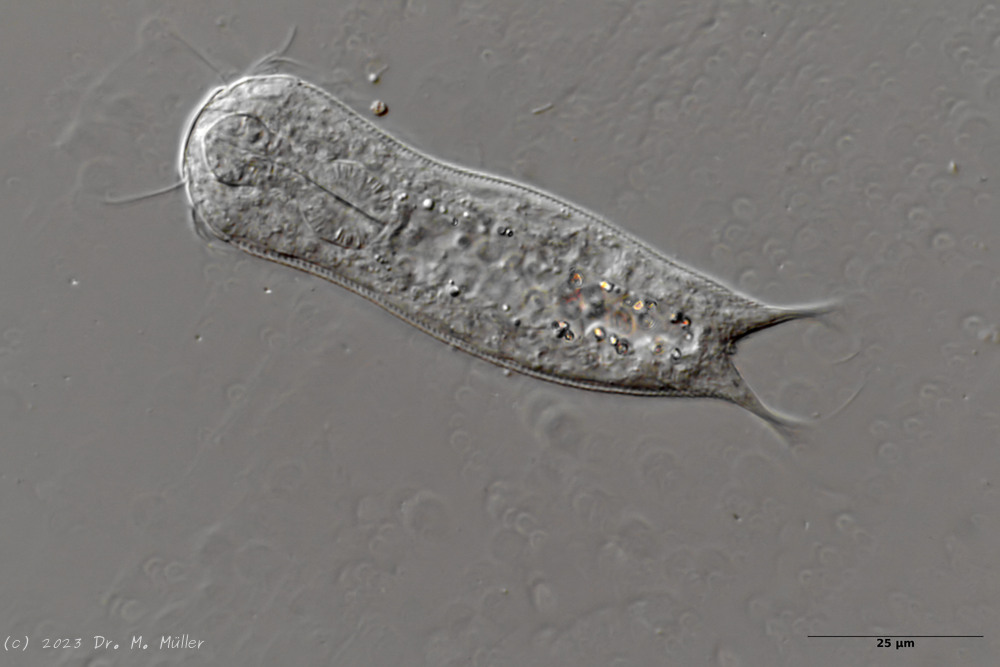
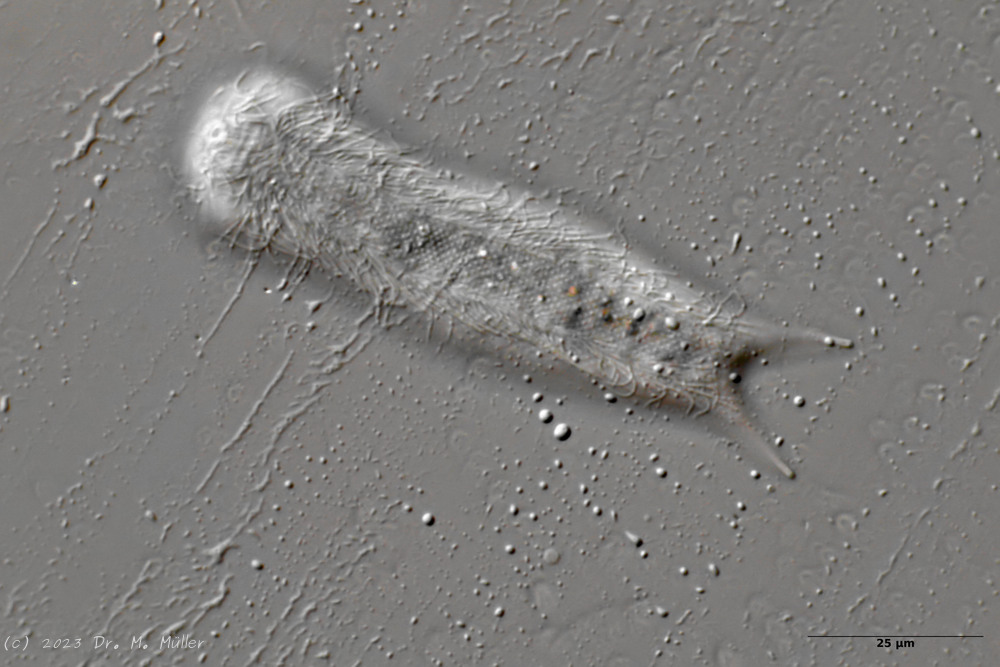

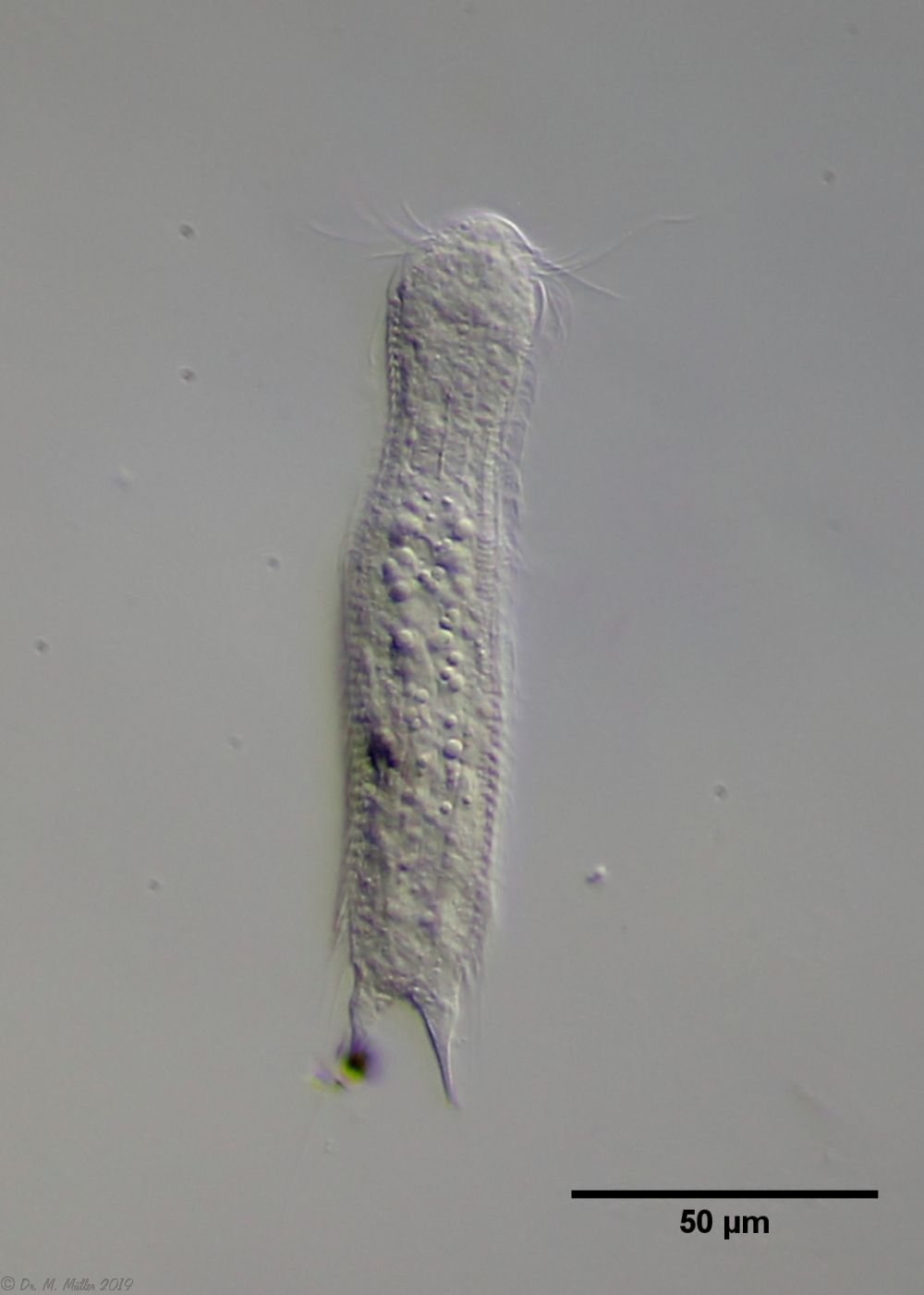
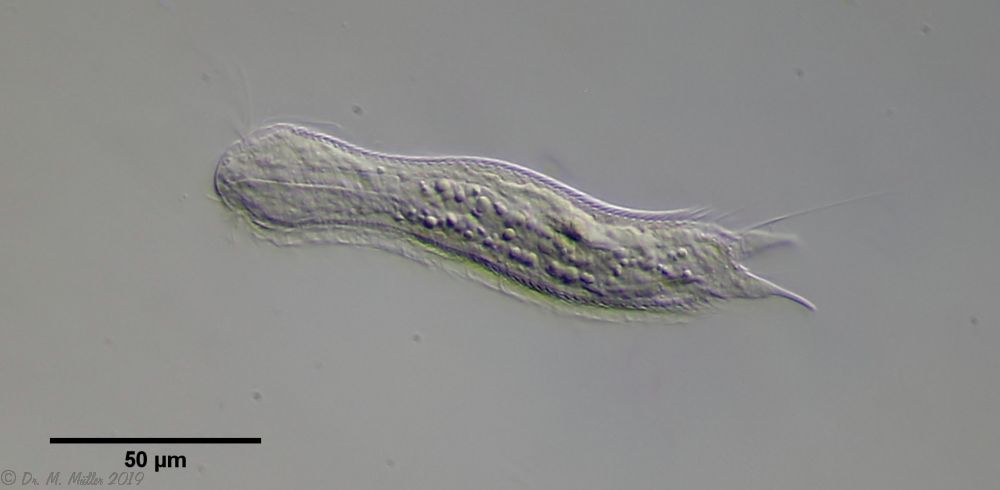


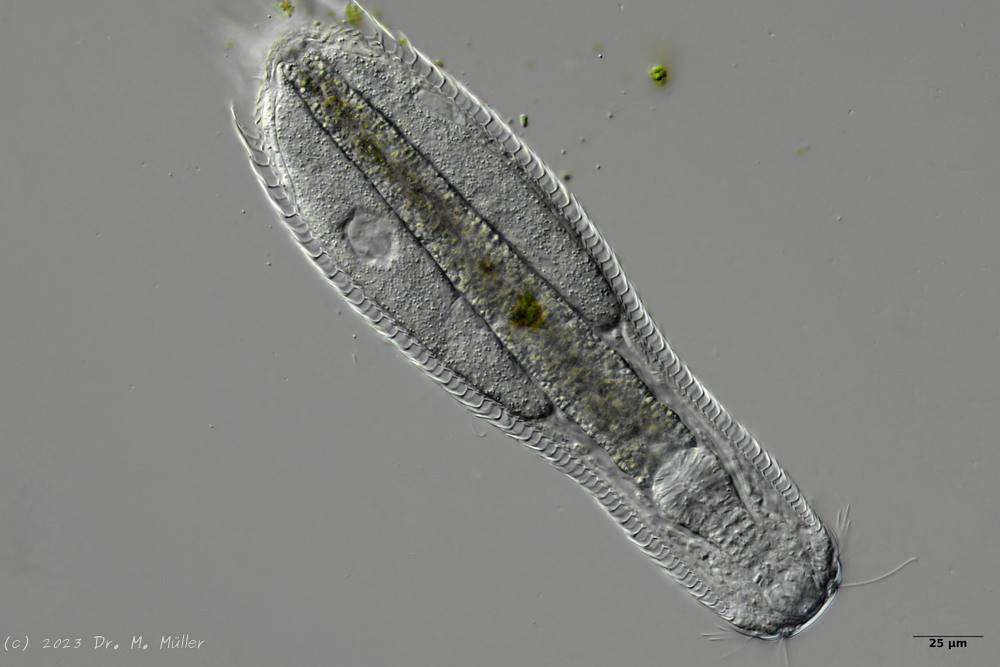

 Backscales
Backscales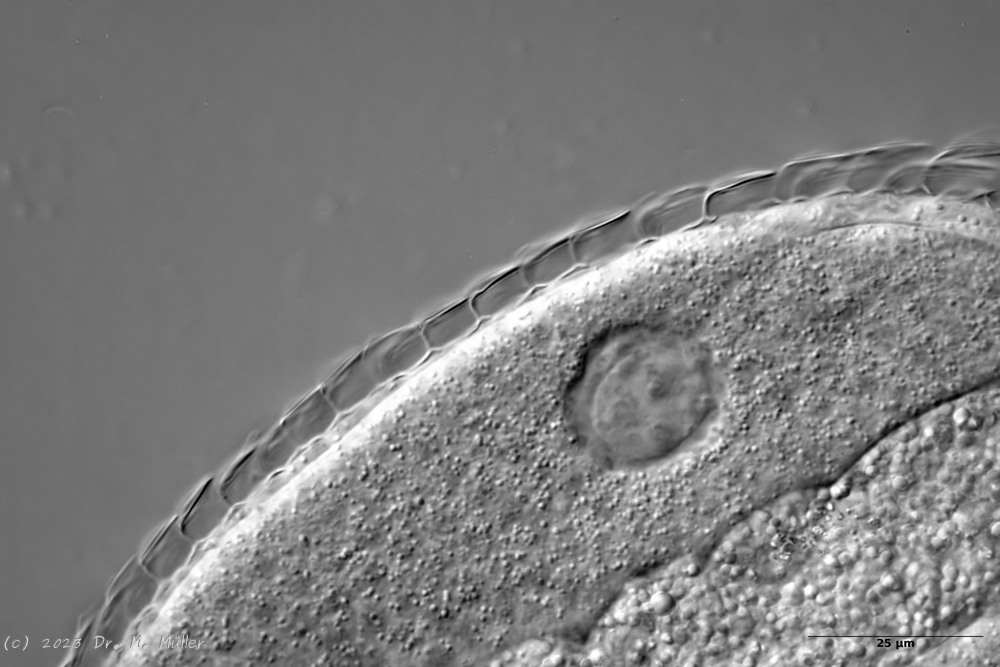
 Cross section scale stems
Cross section scale stems

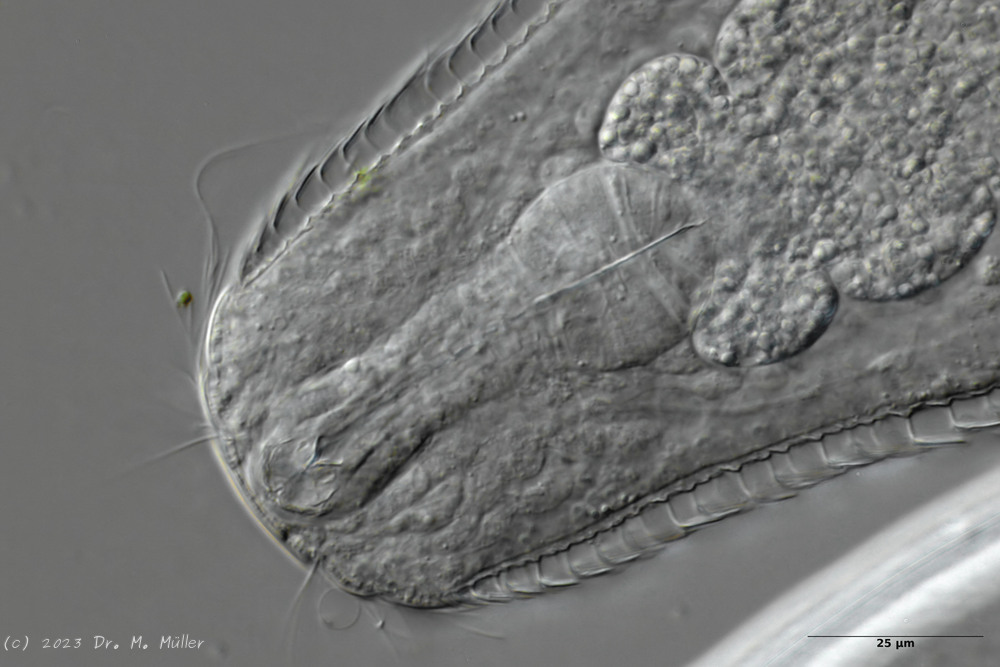

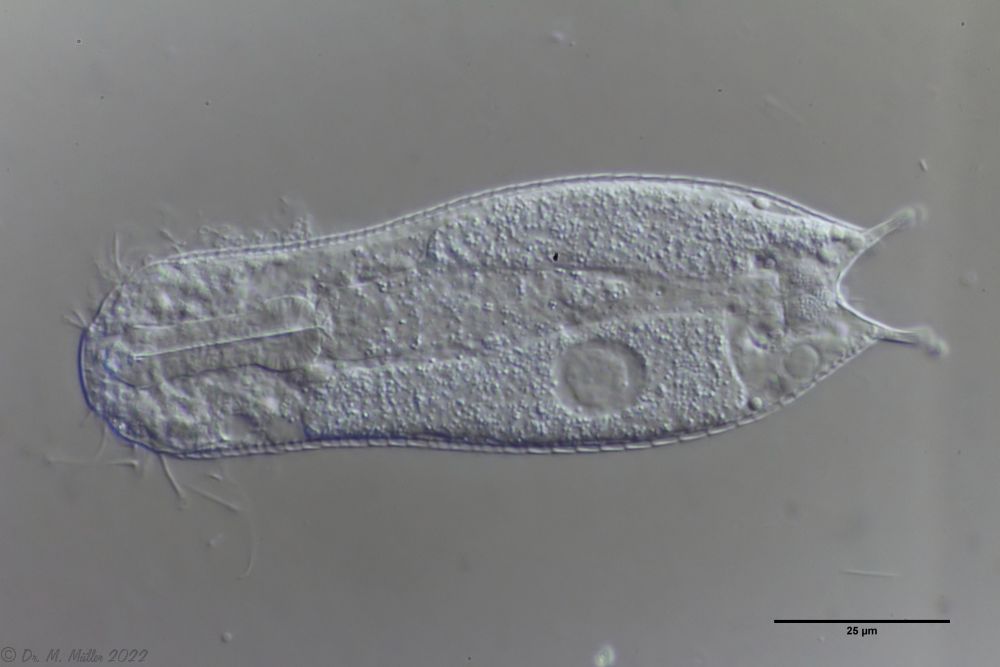
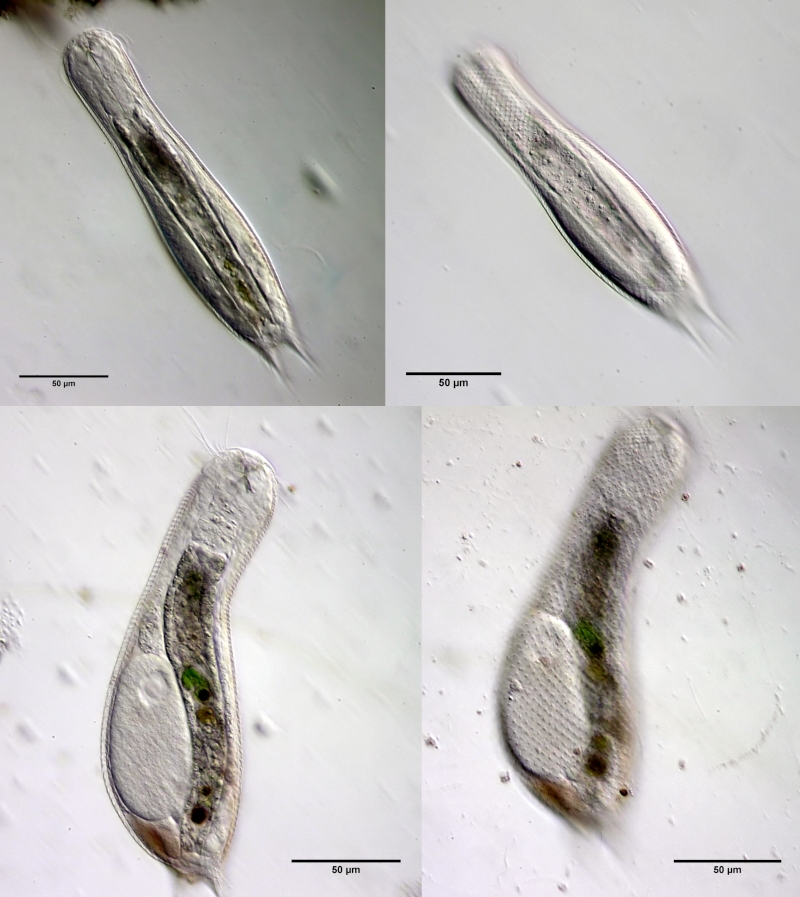
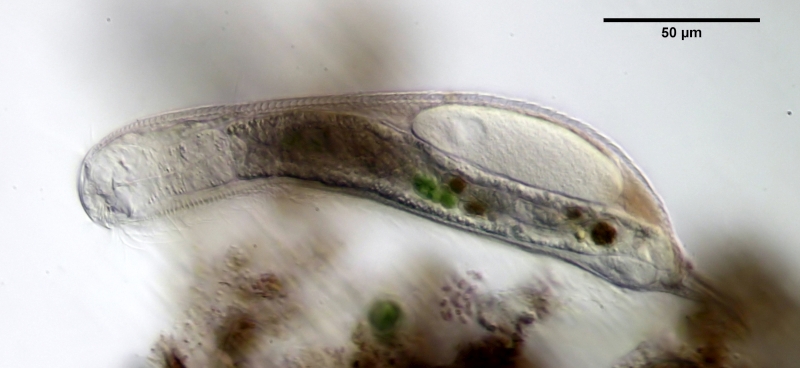

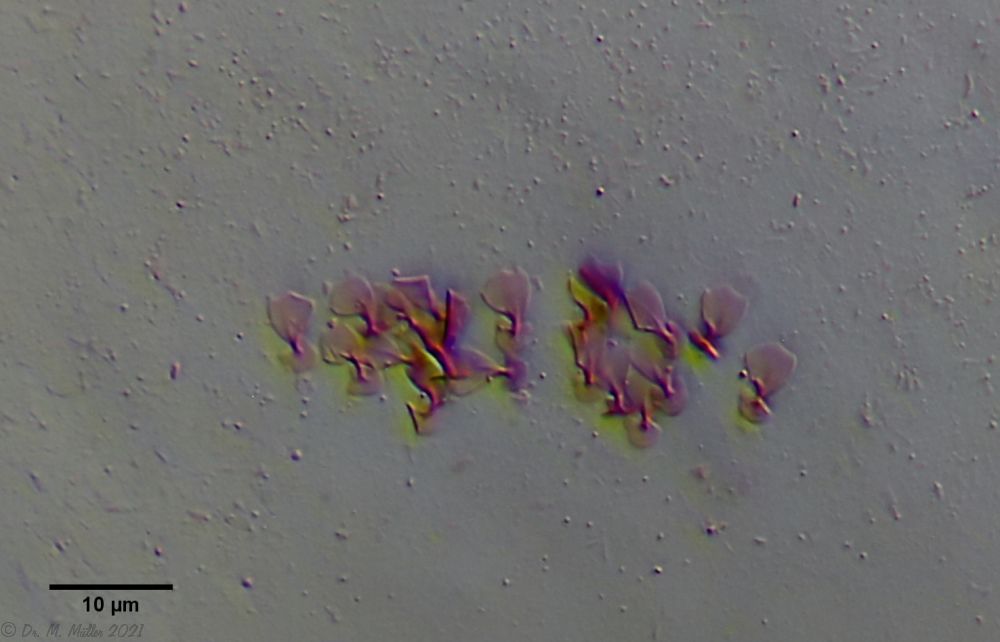
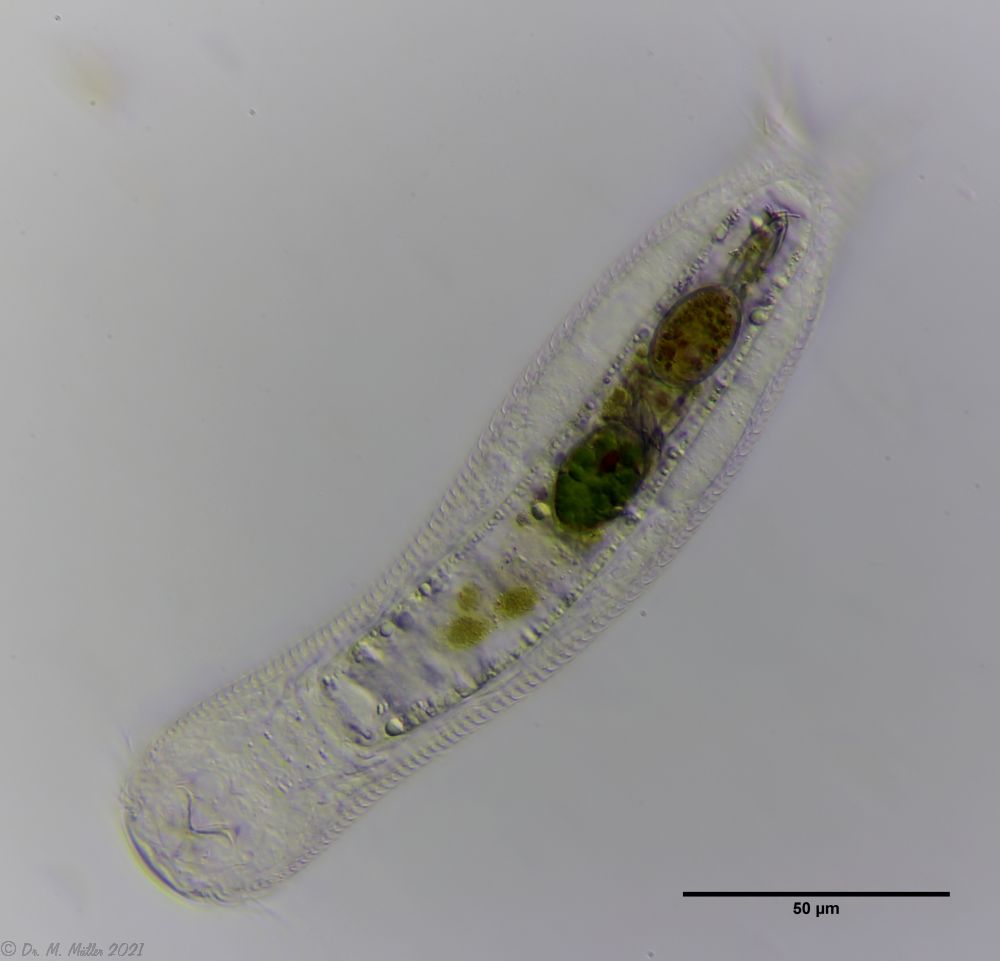
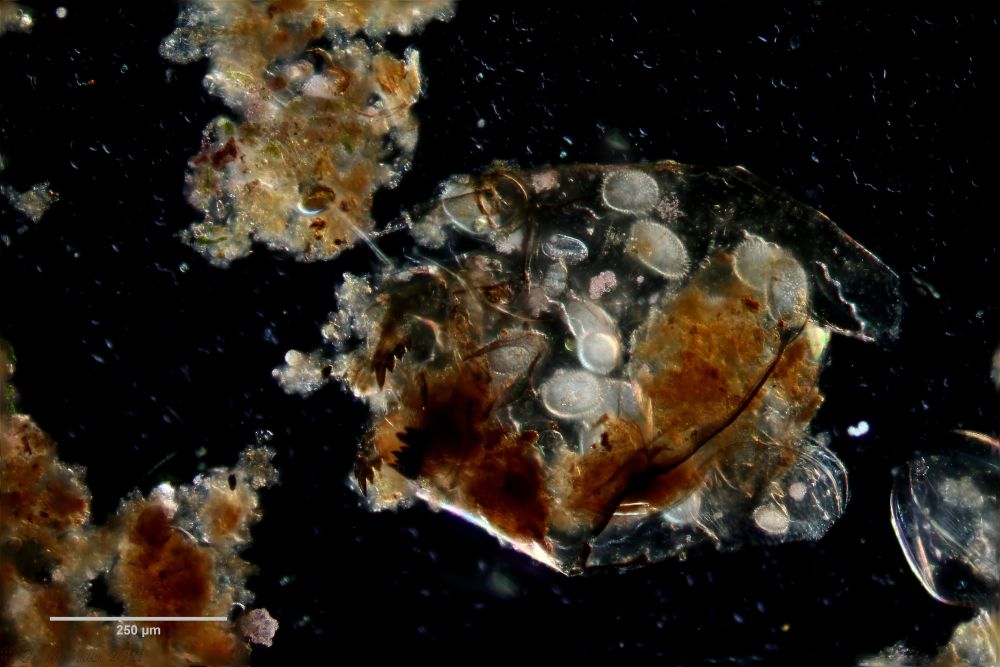
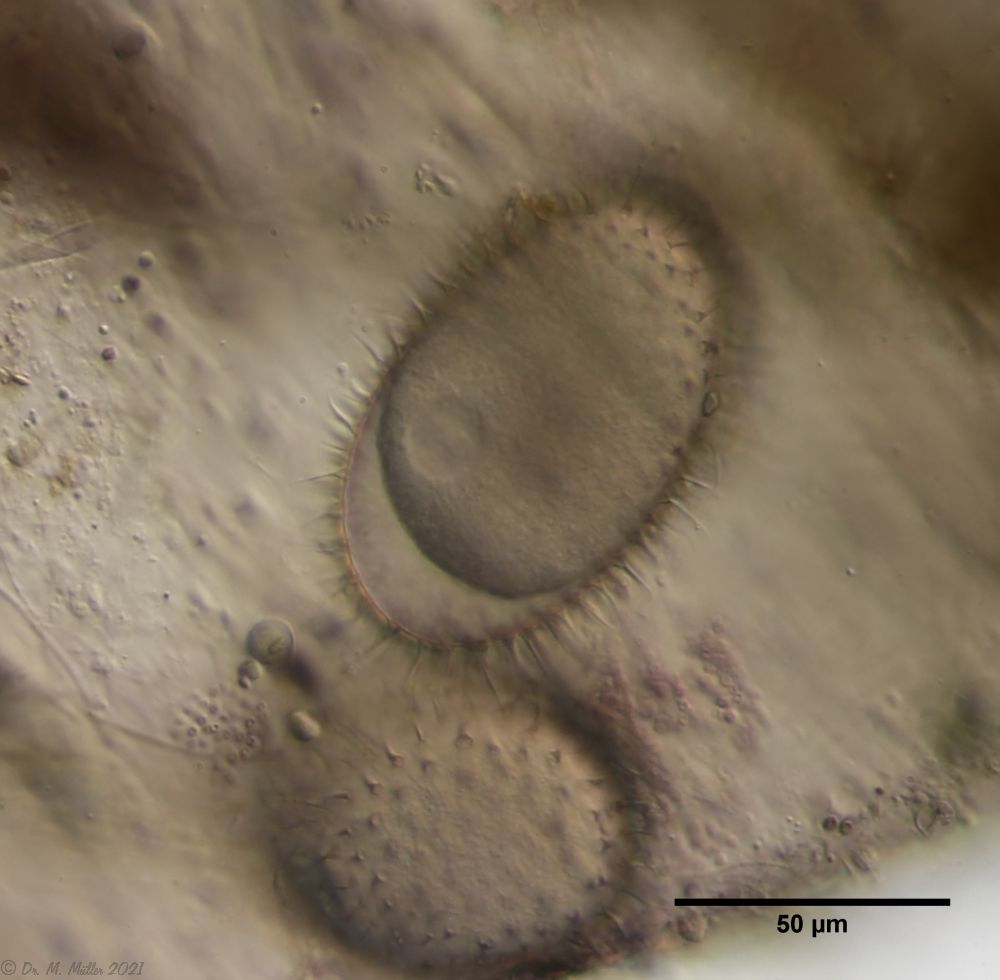

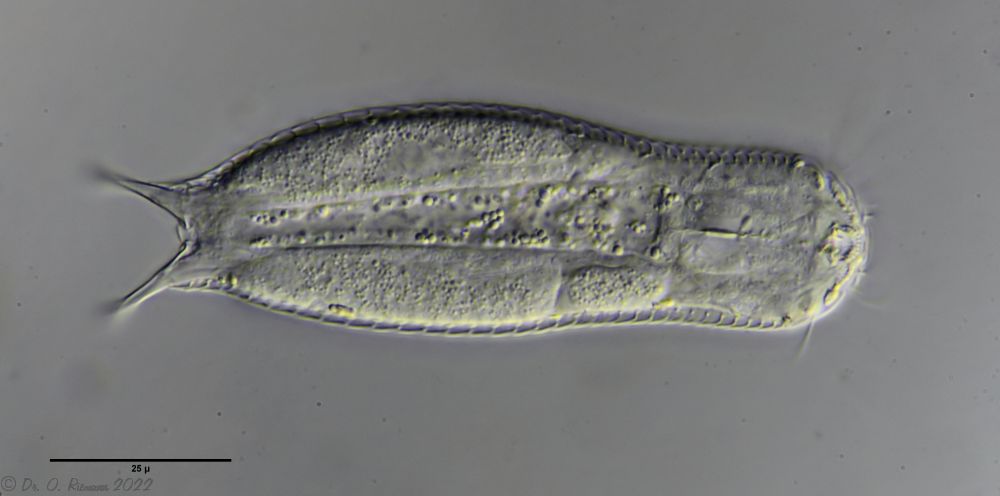

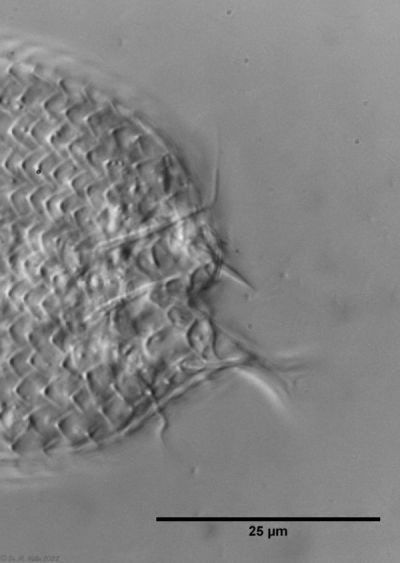

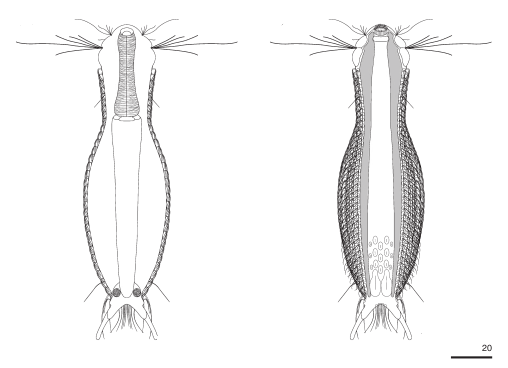
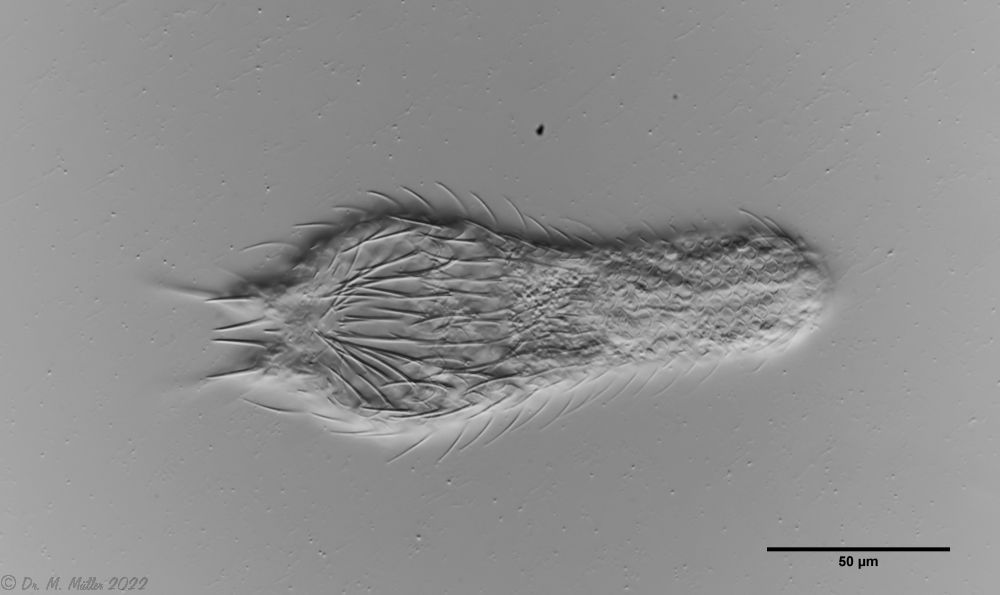
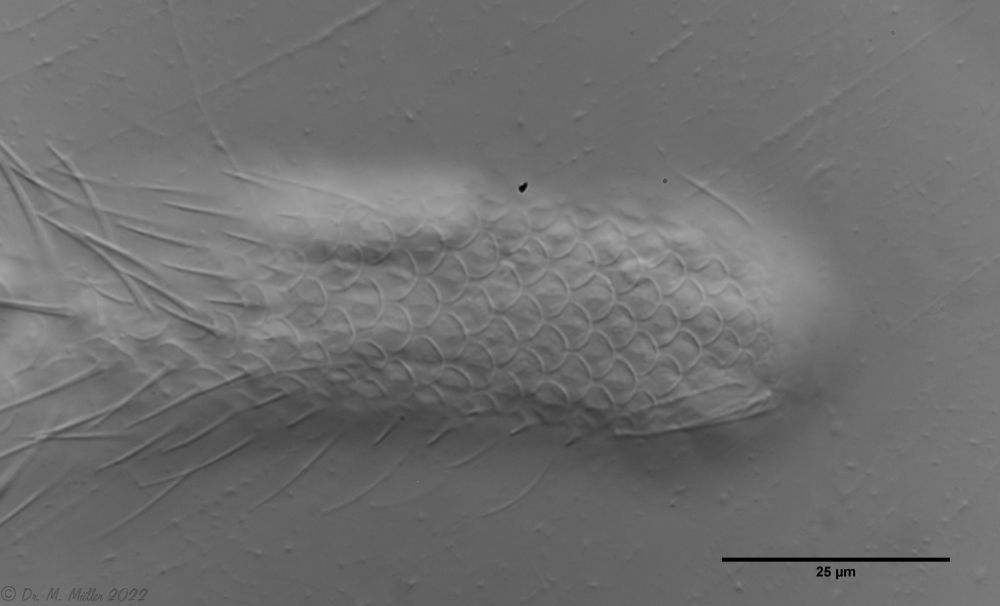
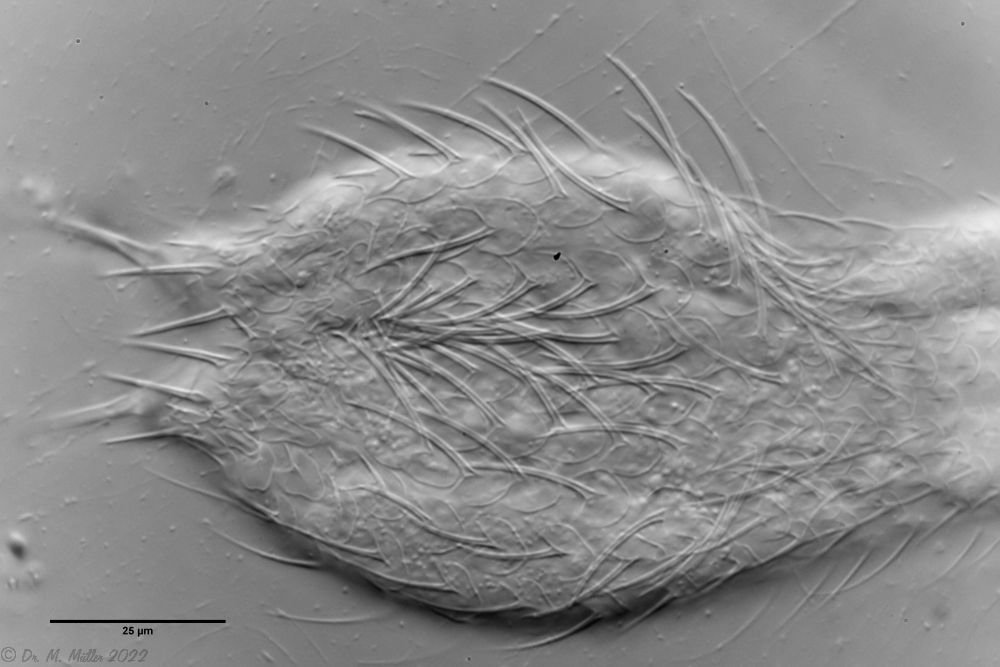

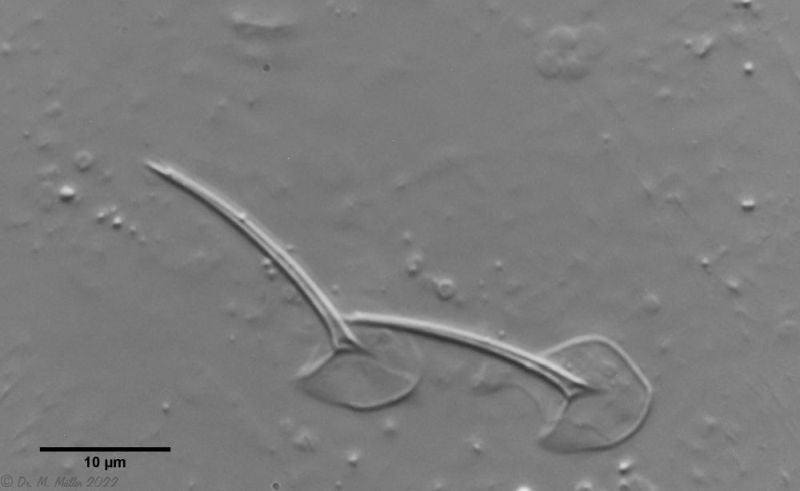
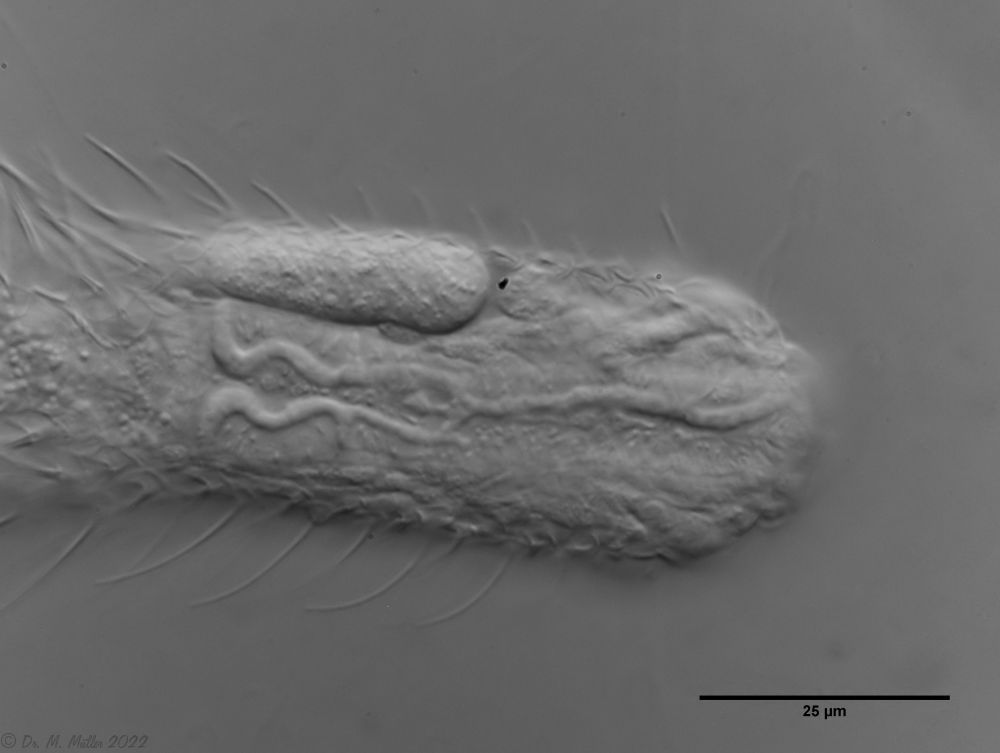
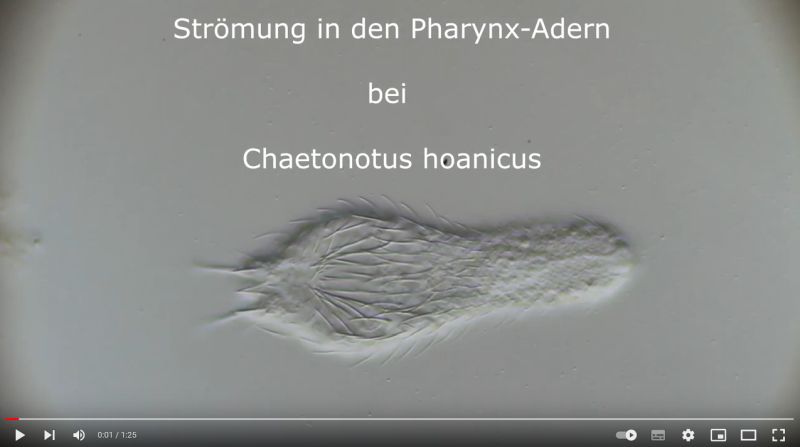
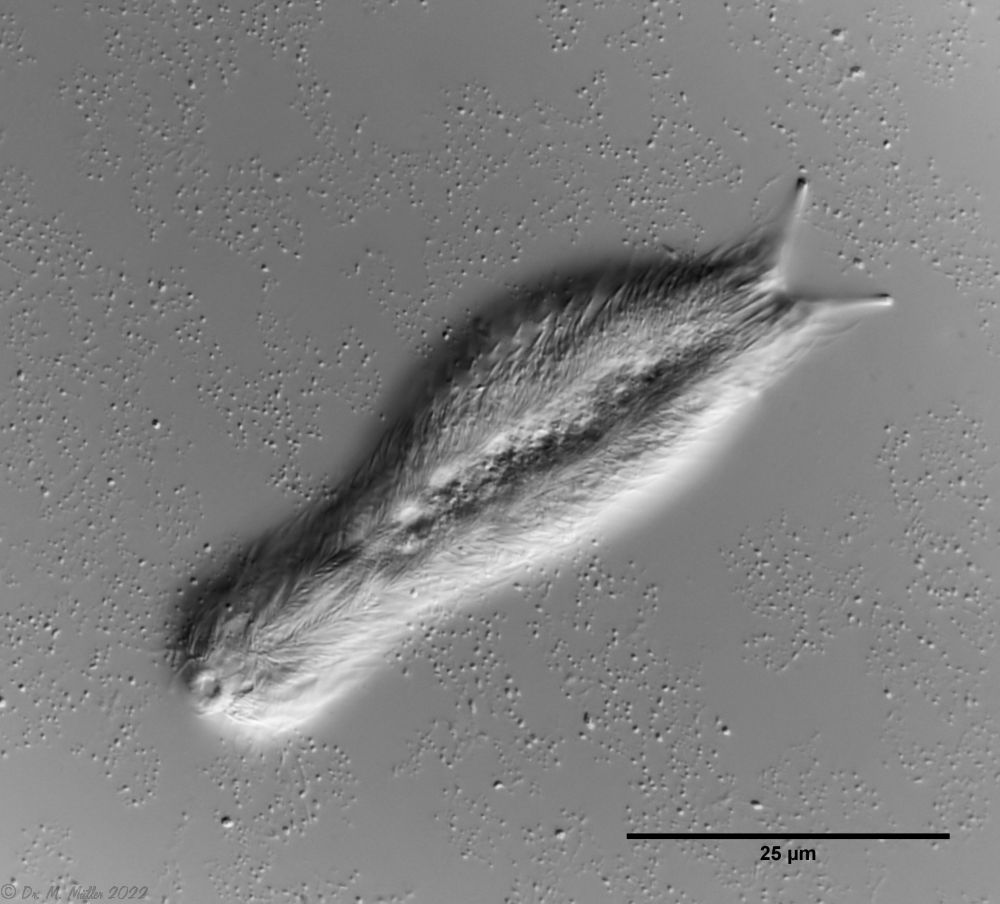 Ch multispinosus: ventral
Ch multispinosus: ventral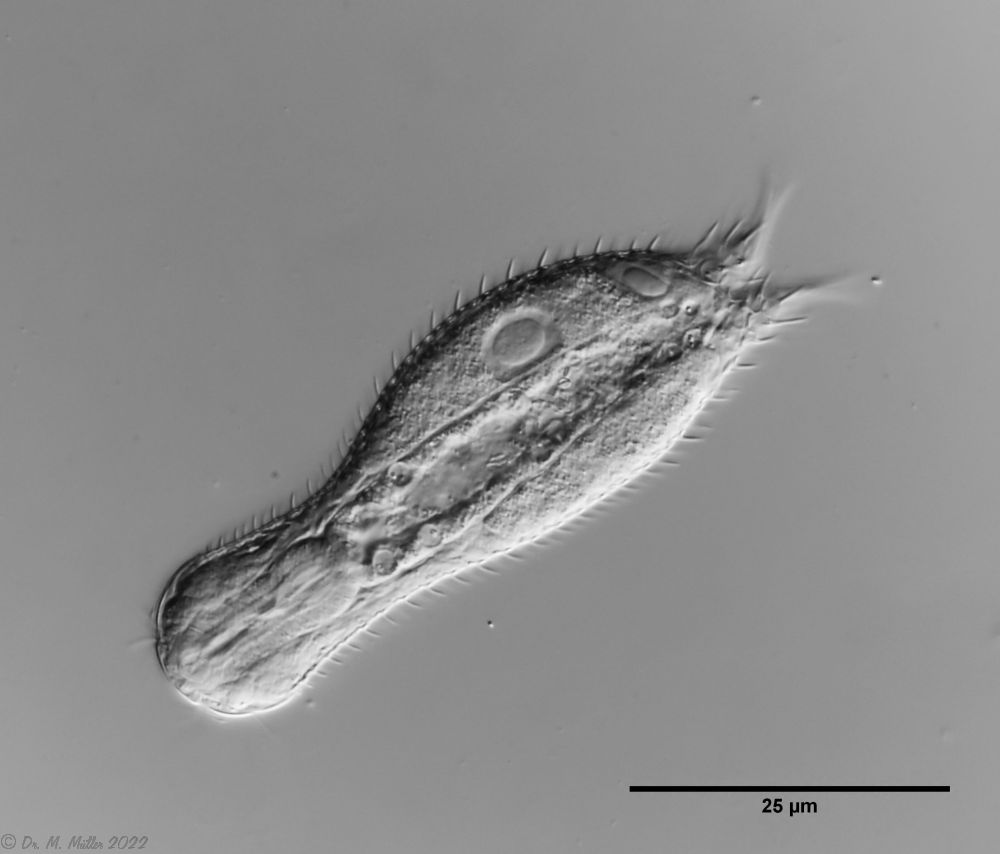 Ch multispinosus: cross section
Ch multispinosus: cross section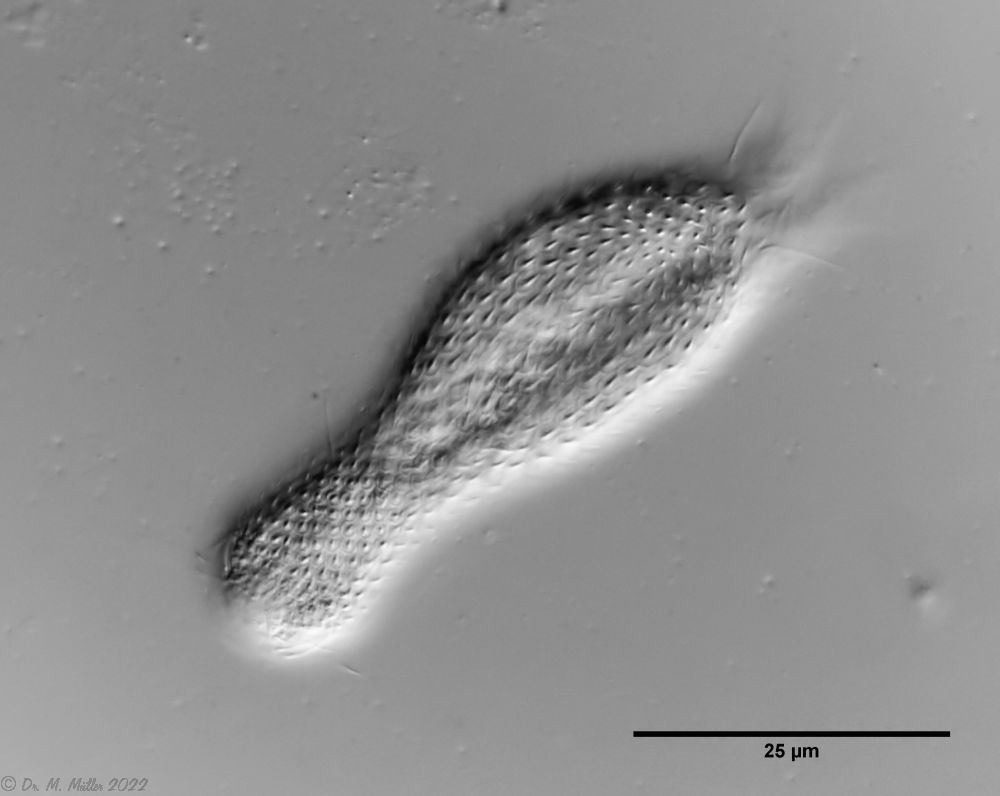 Ch multispinosus: dorsal
Ch multispinosus: dorsal *Typical Chaetonotus (Hystricochaetonotus) - here
*Typical Chaetonotus (Hystricochaetonotus) - here
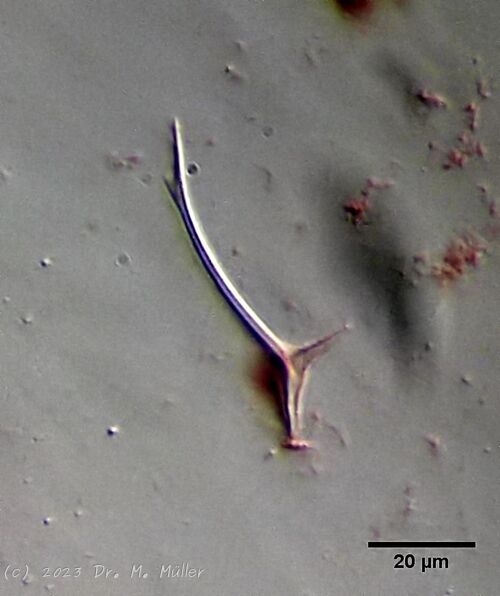
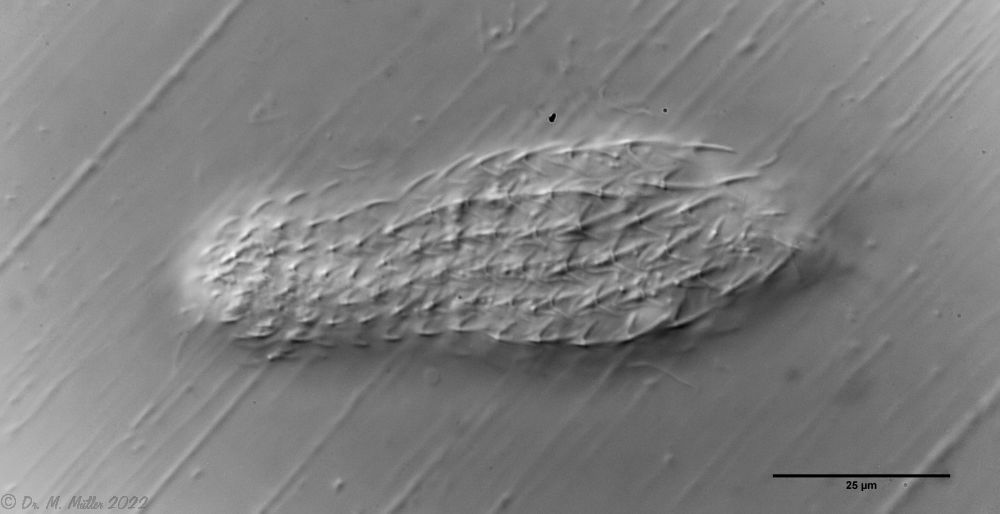
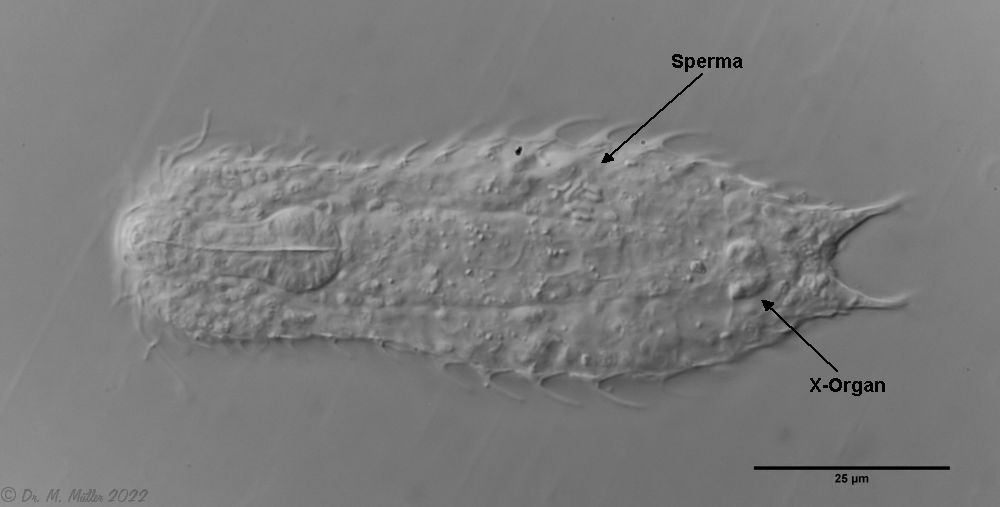
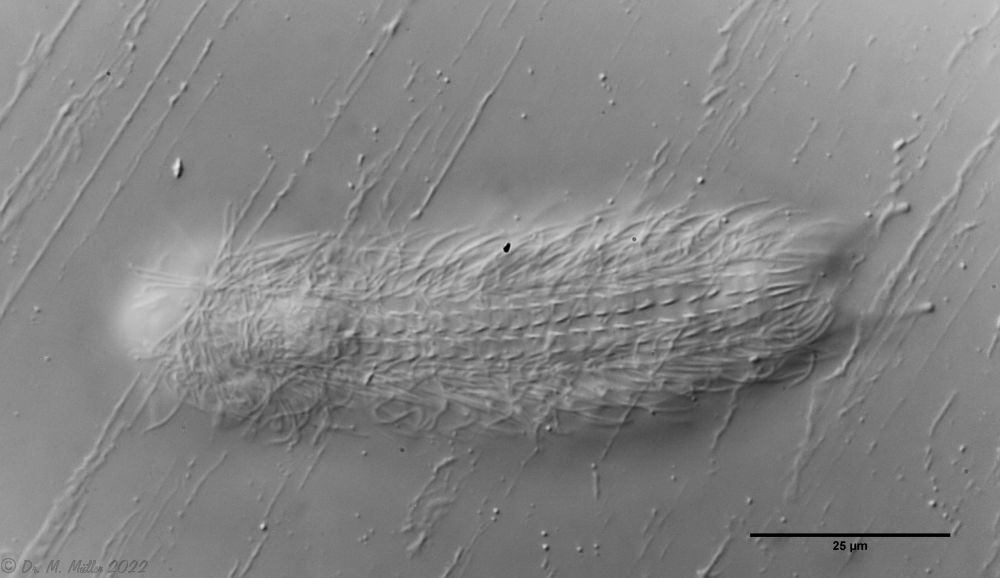
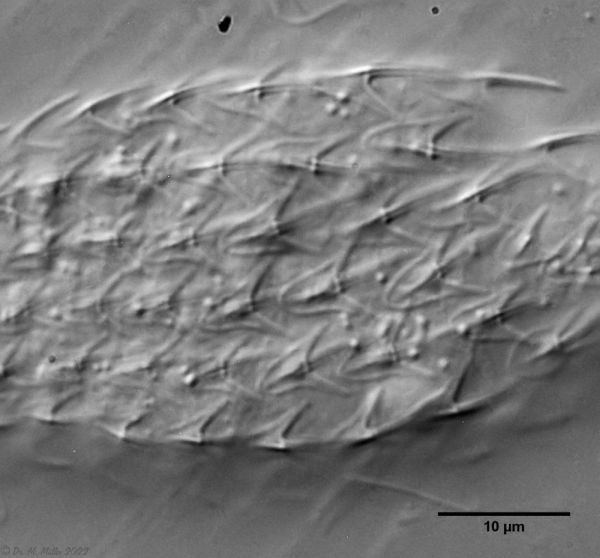
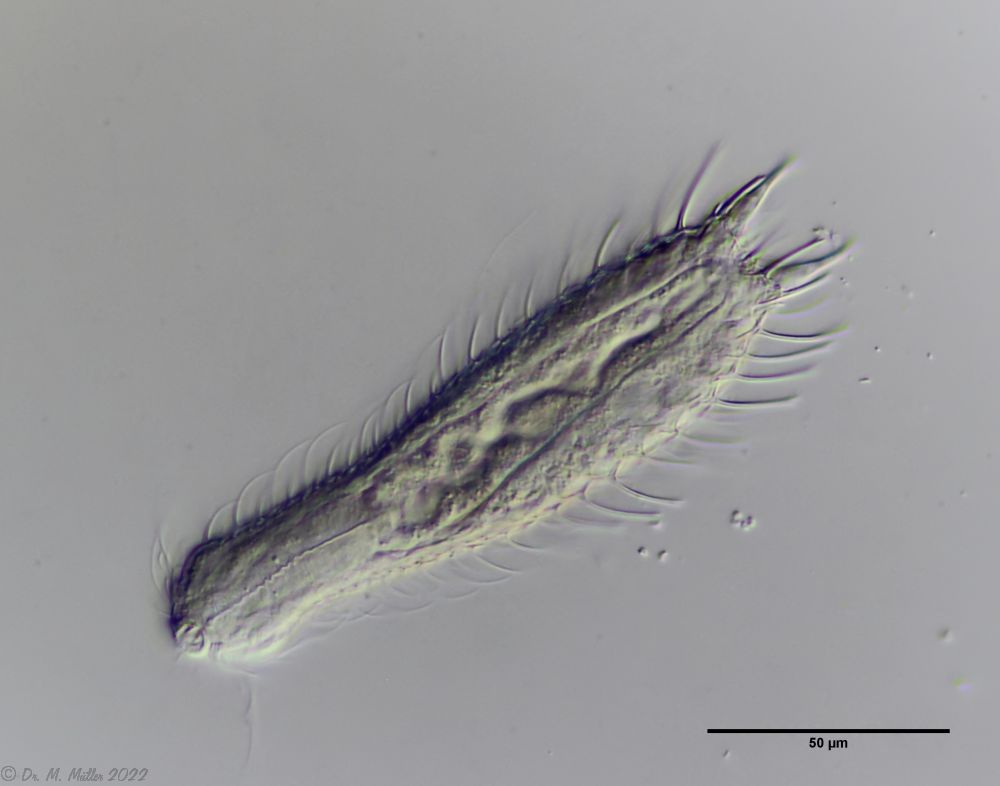 Ch. murrayi: cross section
Ch. murrayi: cross section Ch. murrayi: dorsal
Ch. murrayi: dorsal Ch. murrayi: focus on the spine tips
Ch. murrayi: focus on the spine tips Ch. murrayi: dorsal scaling of the head / neck area
Ch. murrayi: dorsal scaling of the head / neck area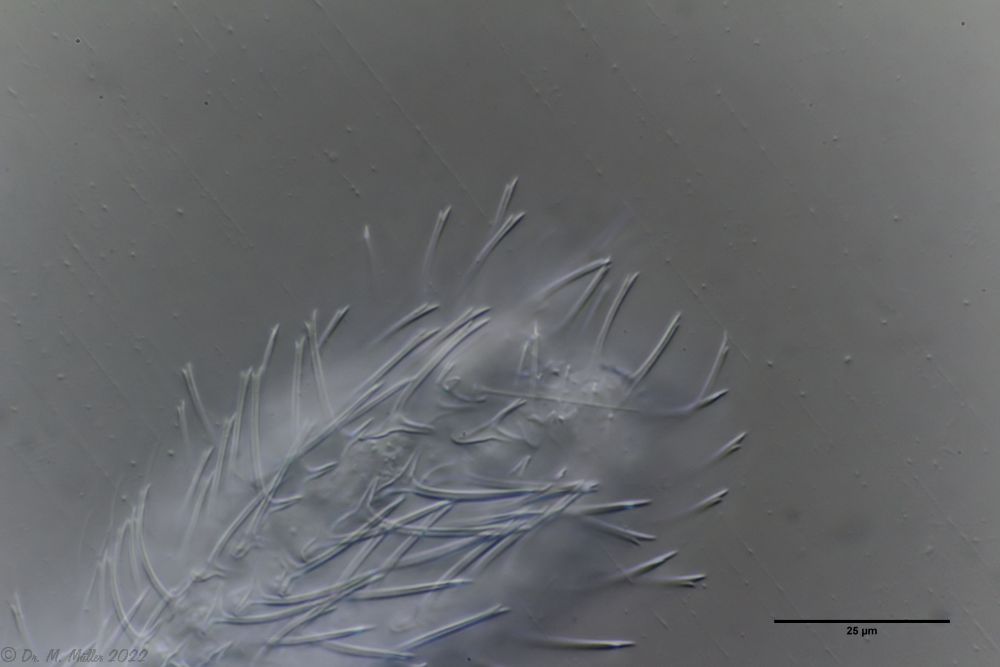 Ch. murrayi: dorsal scaling of the hind end
Ch. murrayi: dorsal scaling of the hind end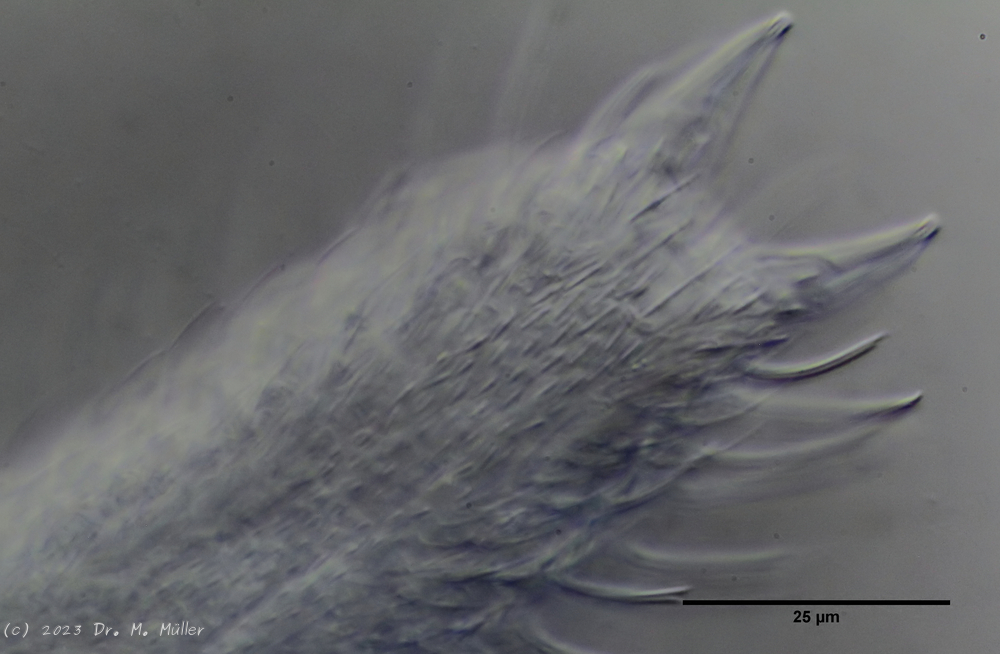 Ch. murray: Scales of the ventral intermediate field
Ch. murray: Scales of the ventral intermediate field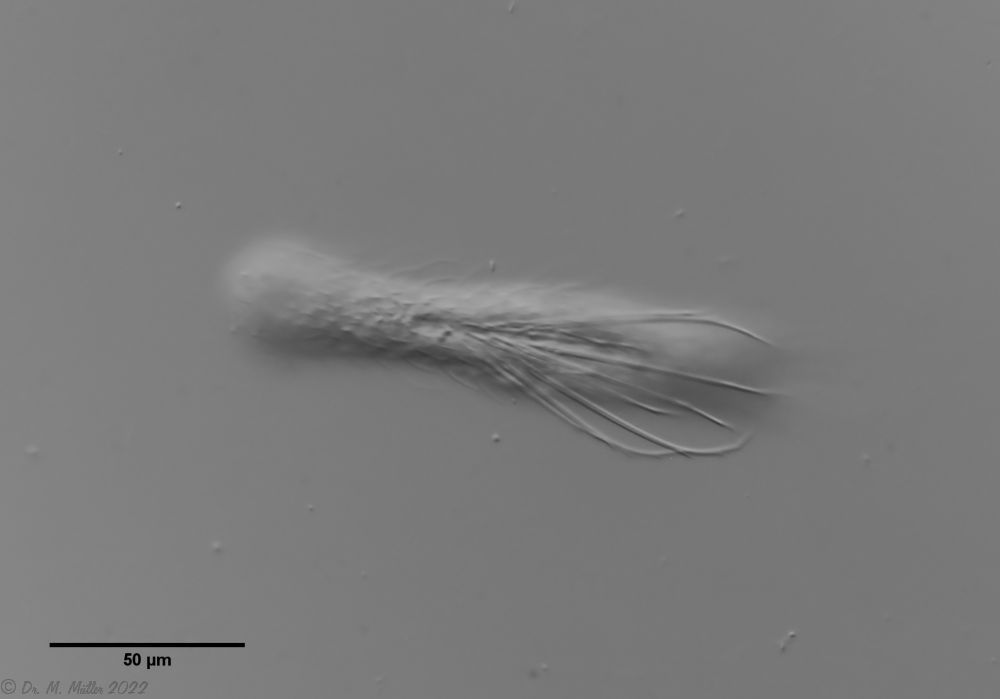 Ch. novenarius: dorsal
Ch. novenarius: dorsal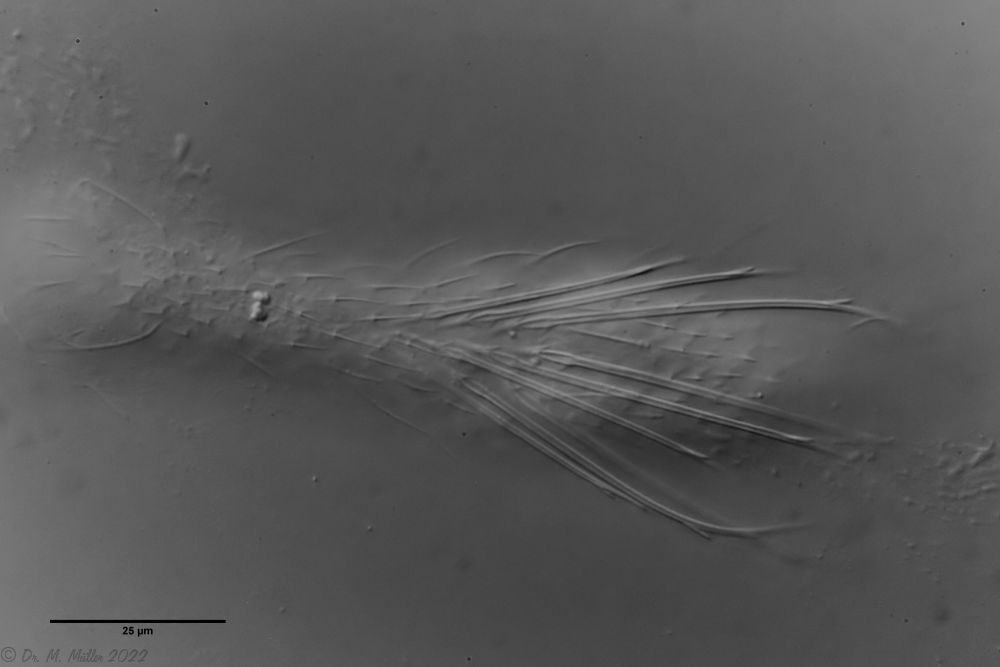 Ch. novenarius: dorsal large spines
Ch. novenarius: dorsal large spines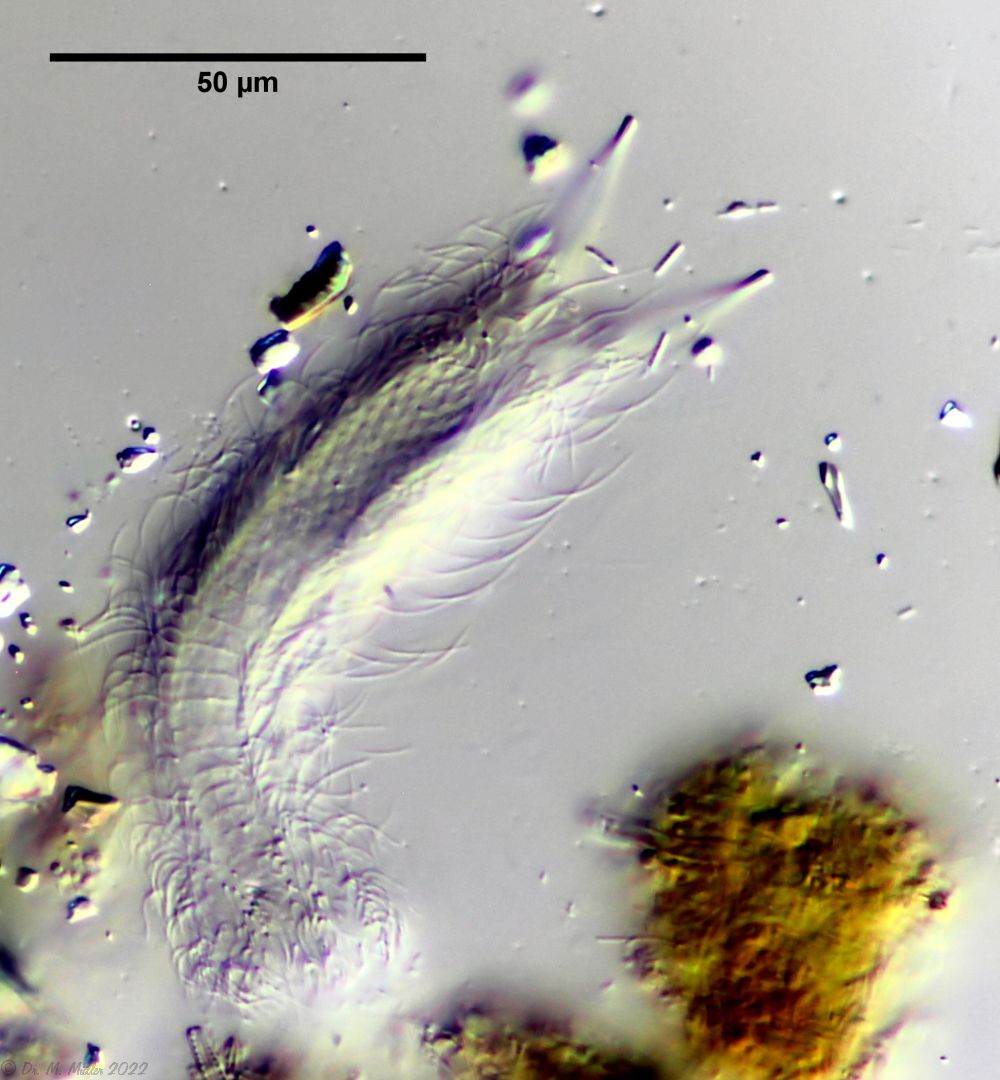 Ch. novenarius: ventral
Ch. novenarius: ventral
 Ch. acanthocephalus: focus on the dorsal scale
Ch. acanthocephalus: focus on the dorsal scale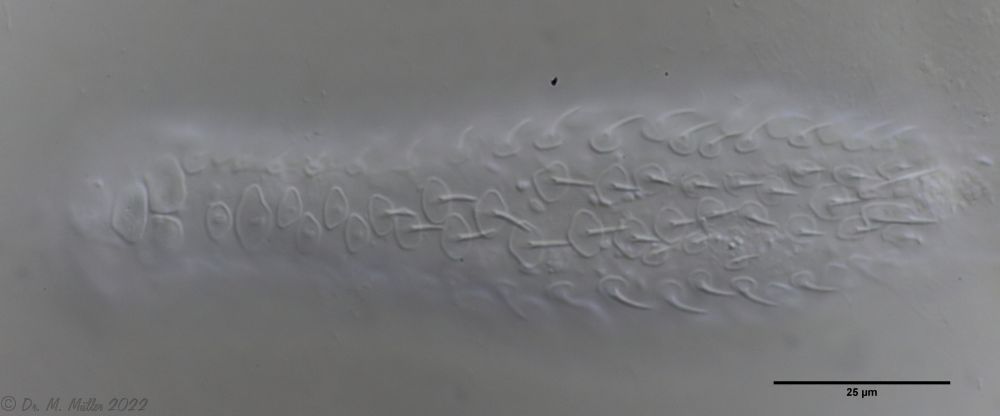 Ch. acanthocephalus: ventral
Ch. acanthocephalus: ventral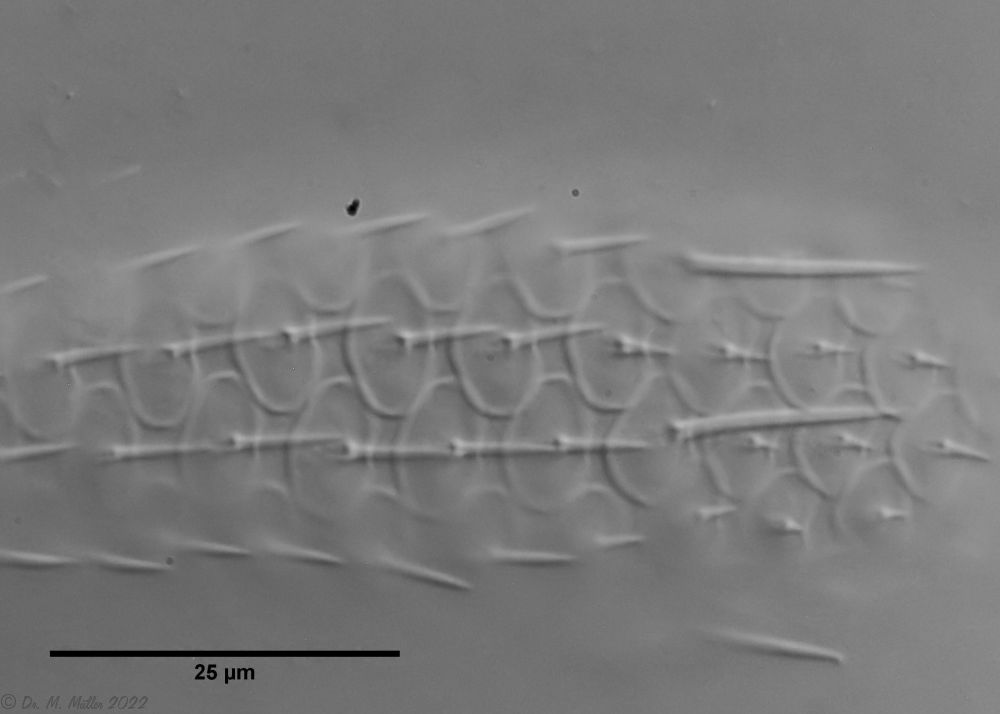 Ch. acanthocephalus: Macerated
Ch. acanthocephalus: Macerated Ch. acanthodes: dorsal scales
Ch. acanthodes: dorsal scales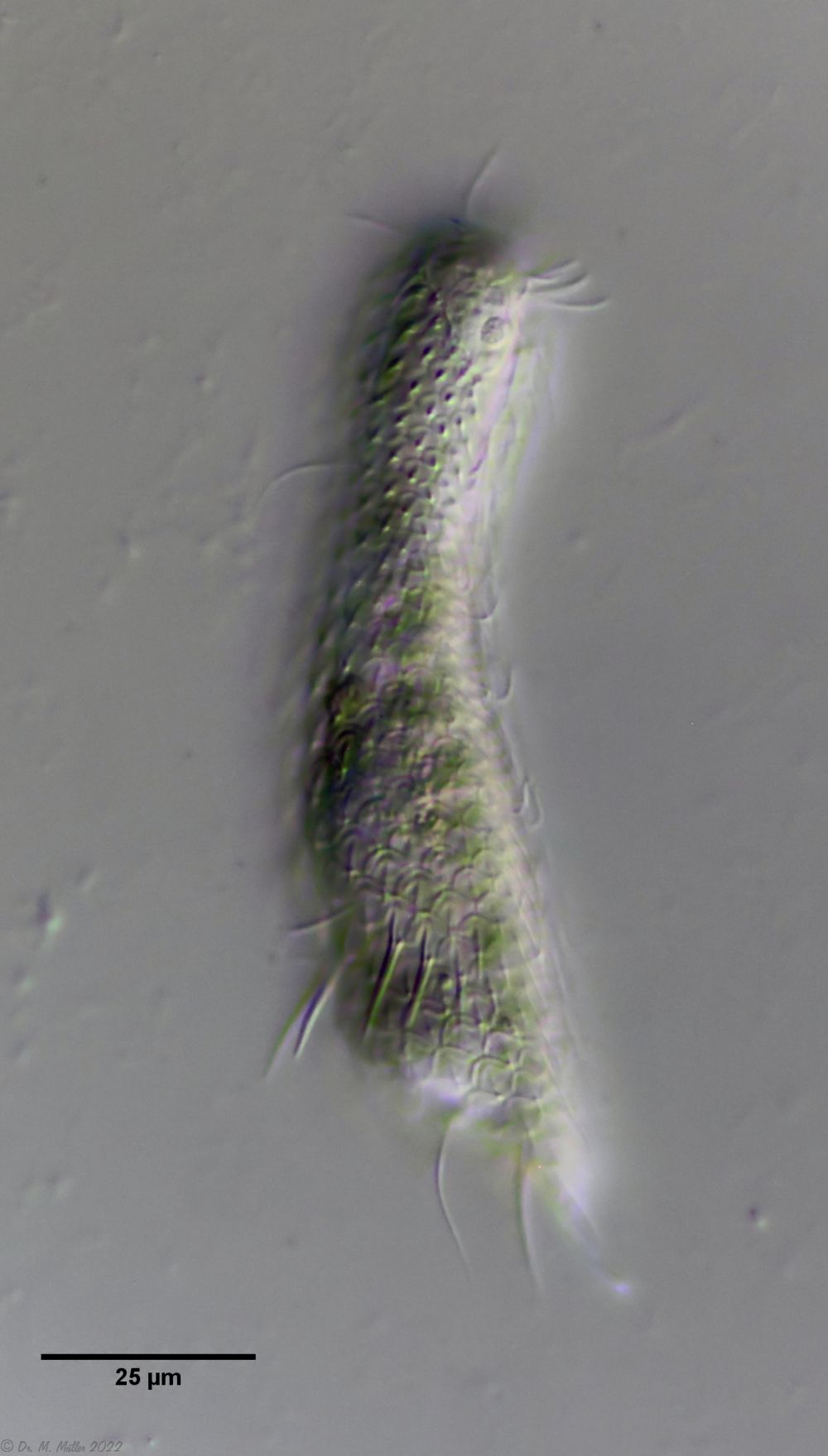 Ch. acanthodes: side view
Ch. acanthodes: side view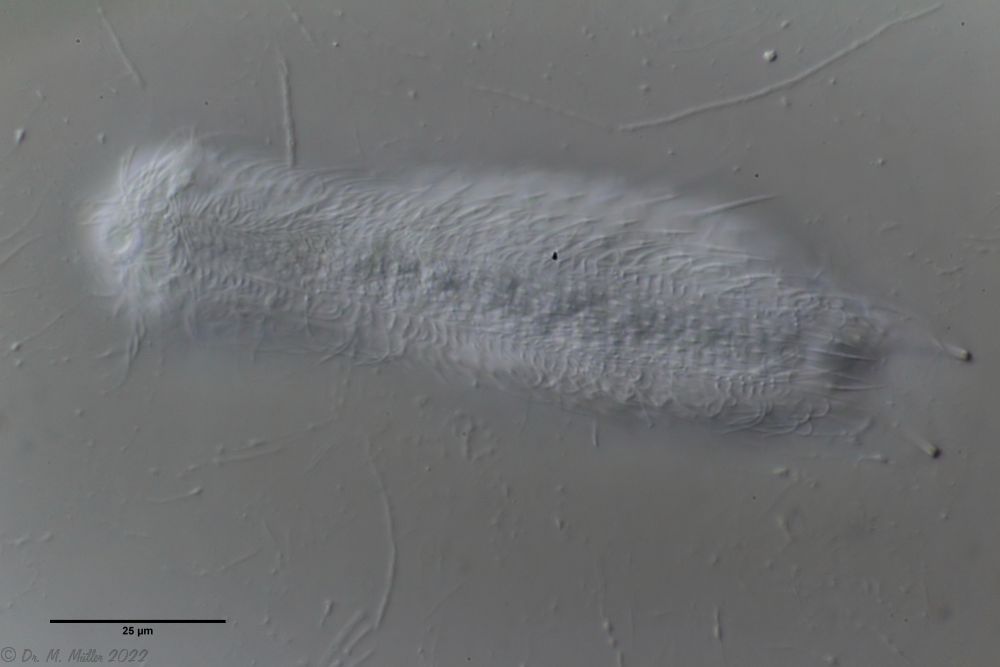 Ch. acanthodes: ventral scales
Ch. acanthodes: ventral scales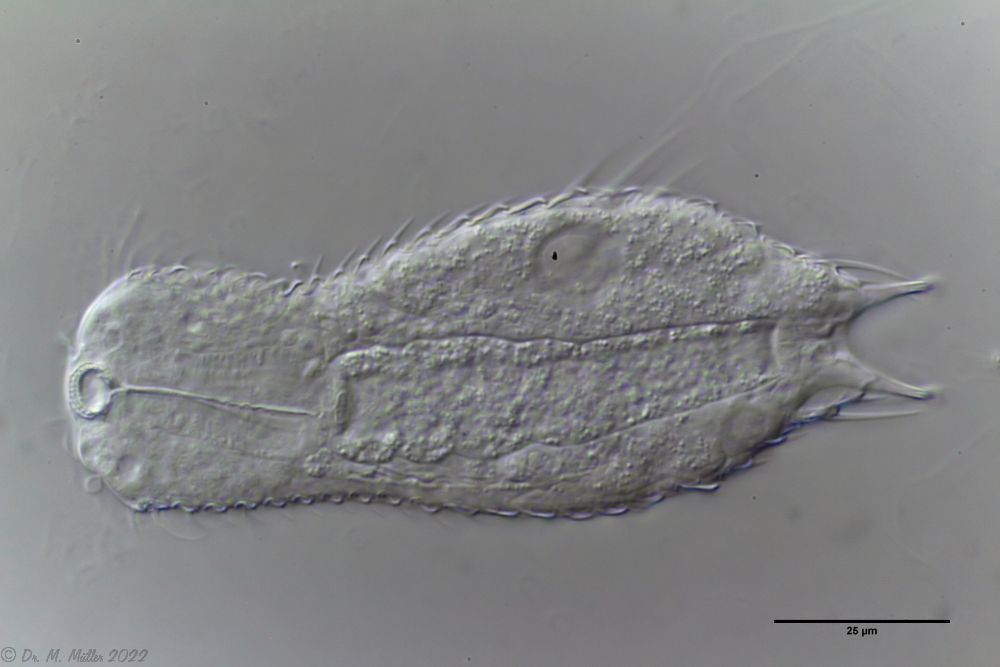 Ch. acanthodes: Cross-section
Ch. acanthodes: Cross-section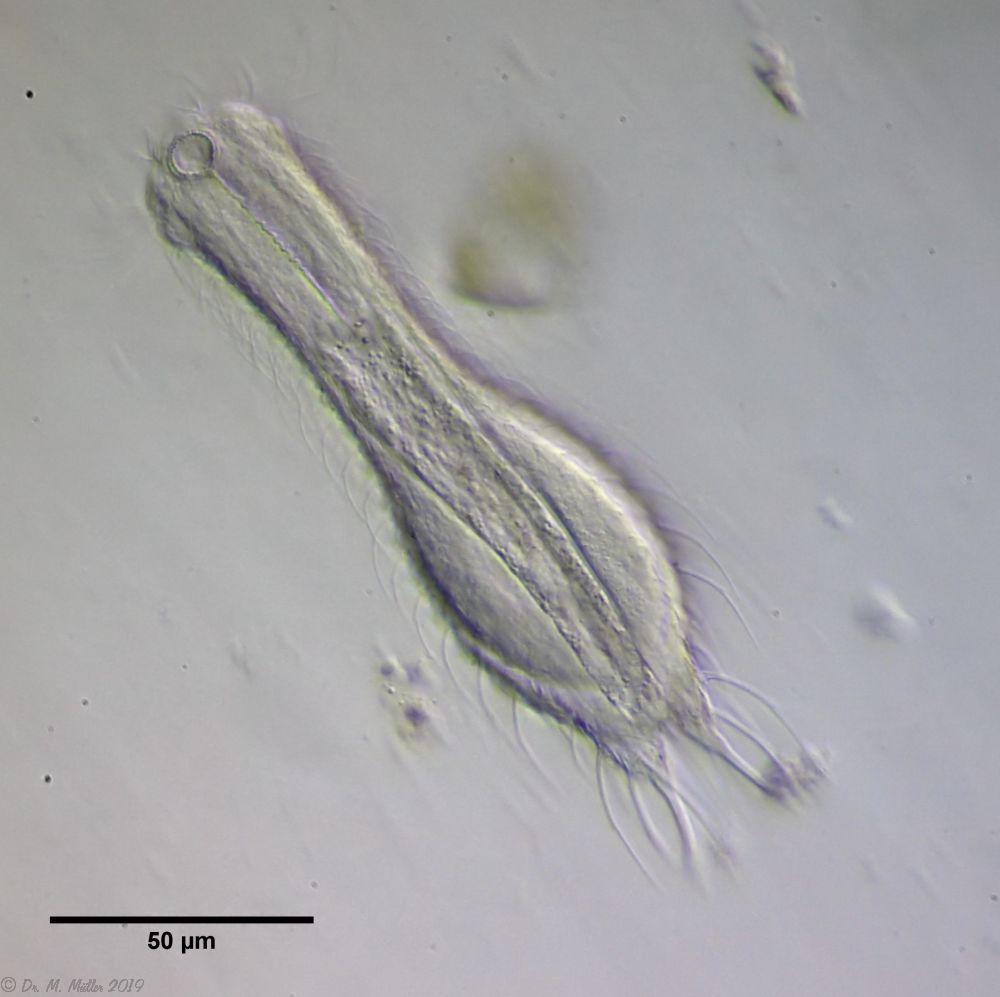 Ch. cordiformis: cross section
Ch. cordiformis: cross section Ch. cordiformis: free swimming specimen with focus on the scale spines
Ch. cordiformis: free swimming specimen with focus on the scale spines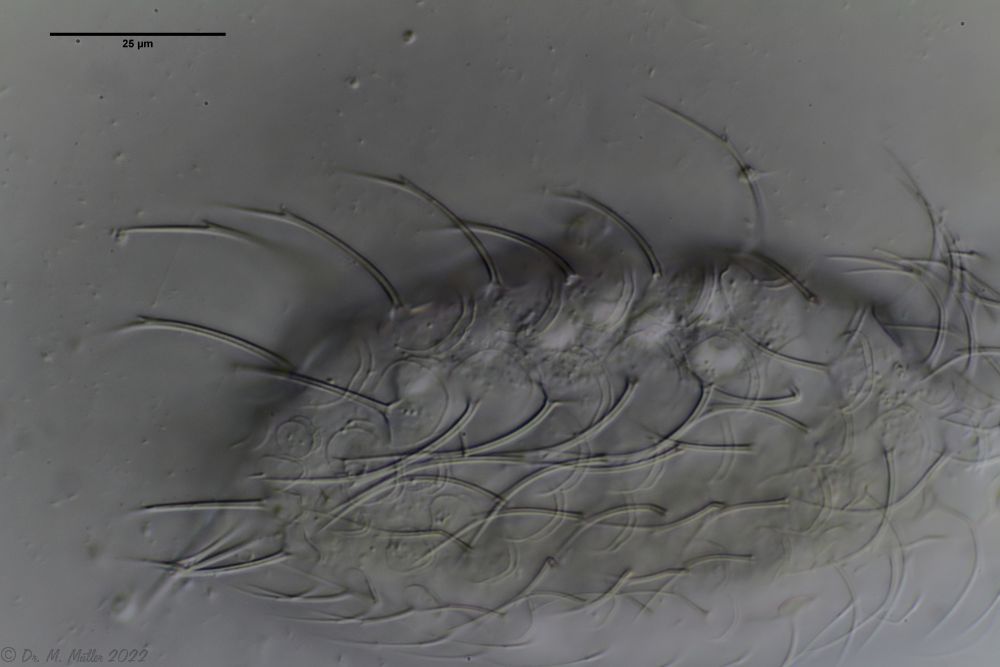 Ch. cordiformis: spiny scales with secondary tip.
Ch. cordiformis: spiny scales with secondary tip.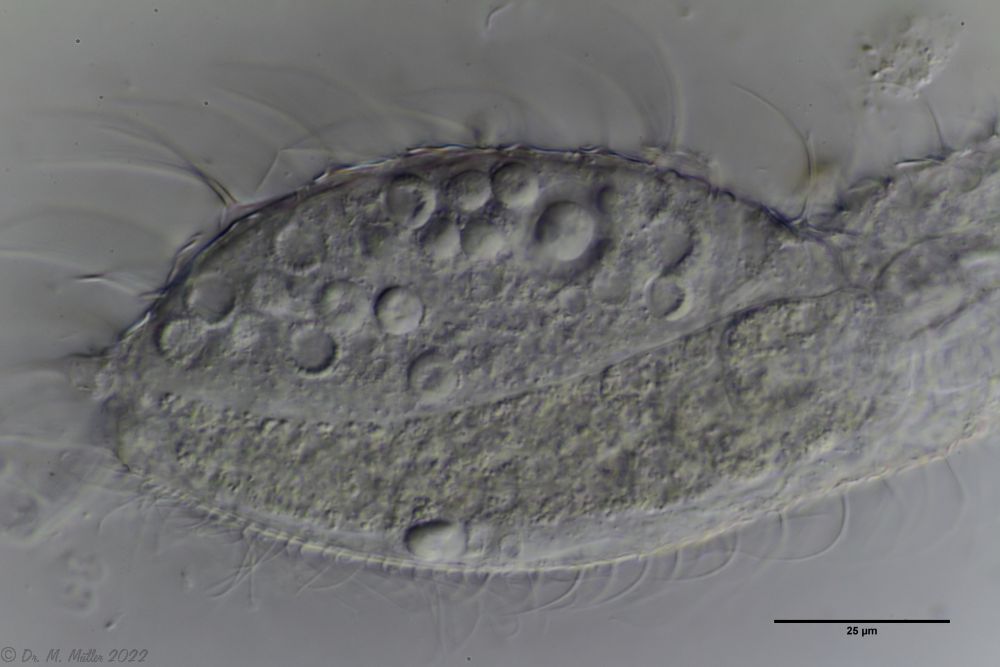 Ch. cordiformis: spherical inclusions.
Ch. cordiformis: spherical inclusions. Ch. cordiformis: heart-shaped scales
Ch. cordiformis: heart-shaped scales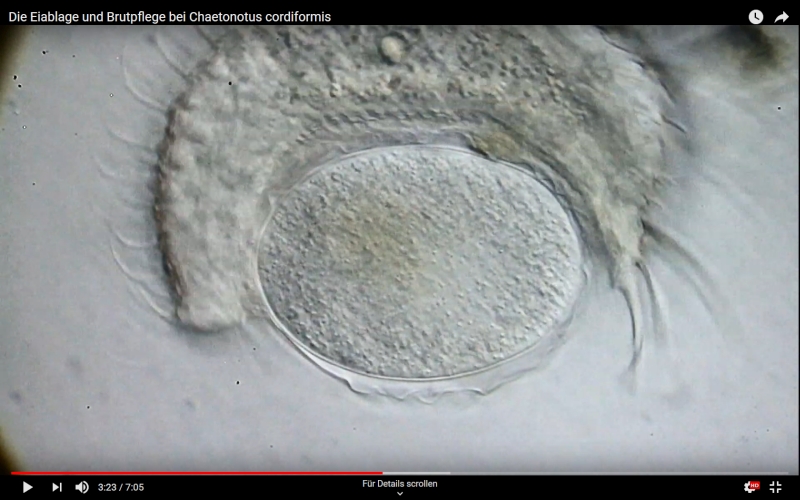
 Dorsal; ds: dorsal spines (5 pairs).
Dorsal; ds: dorsal spines (5 pairs).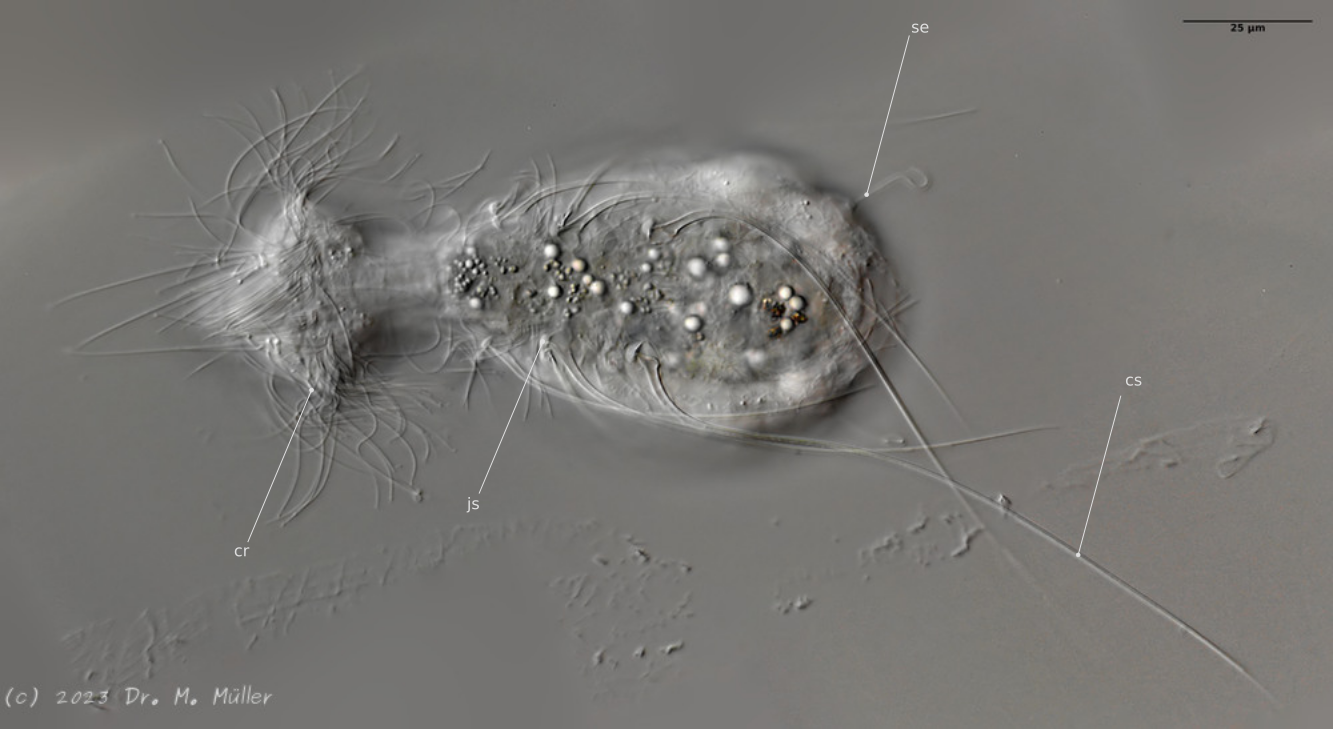
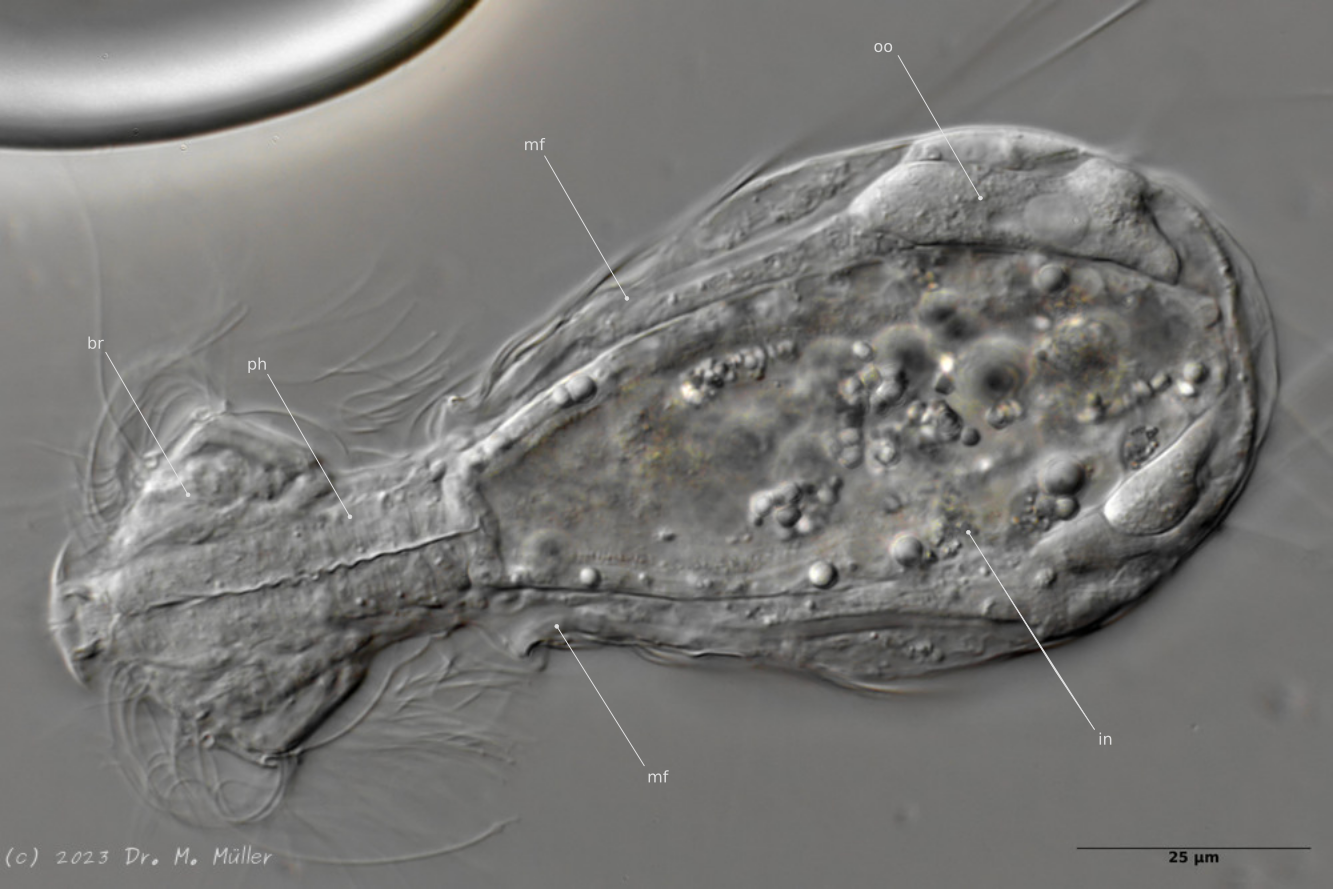
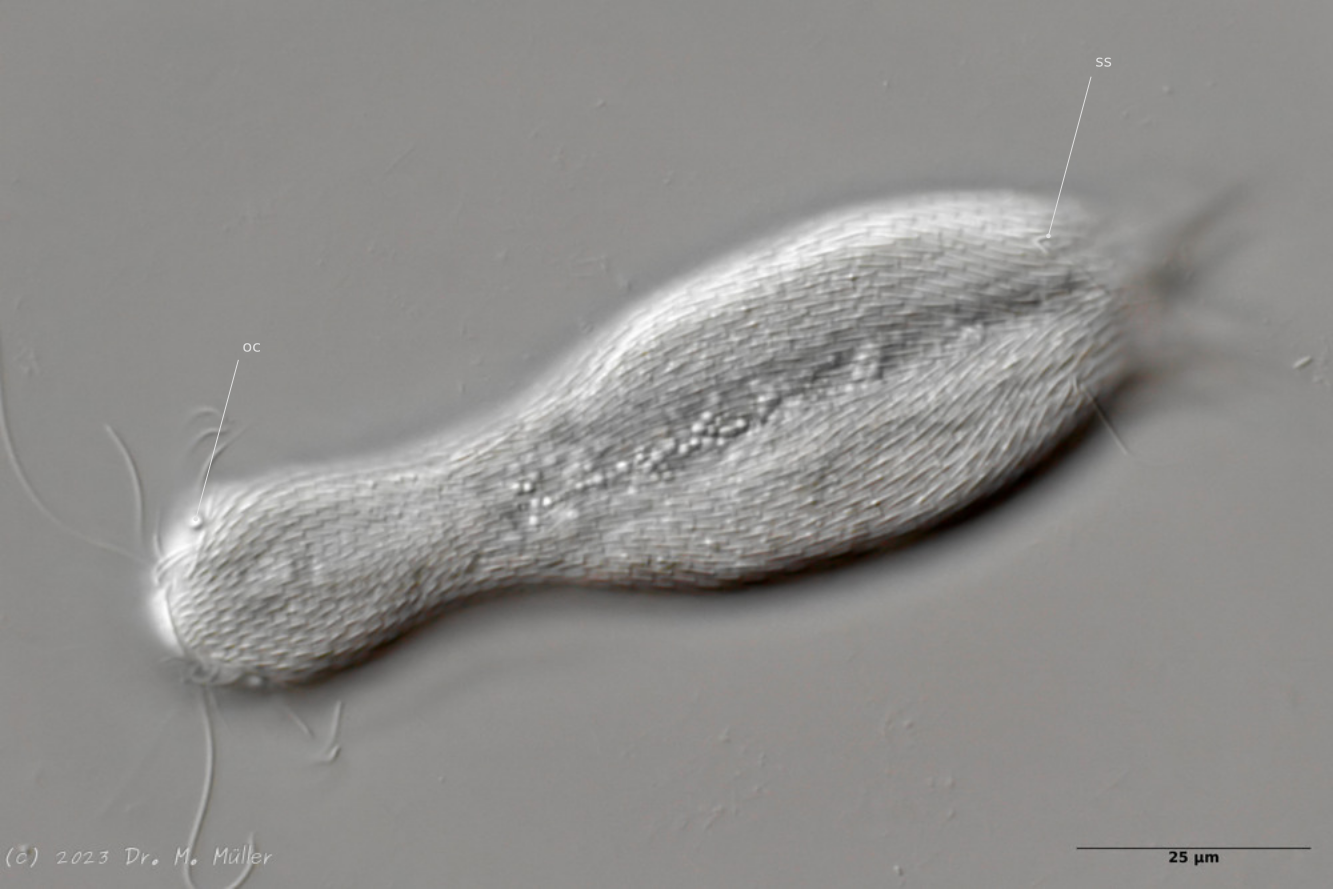
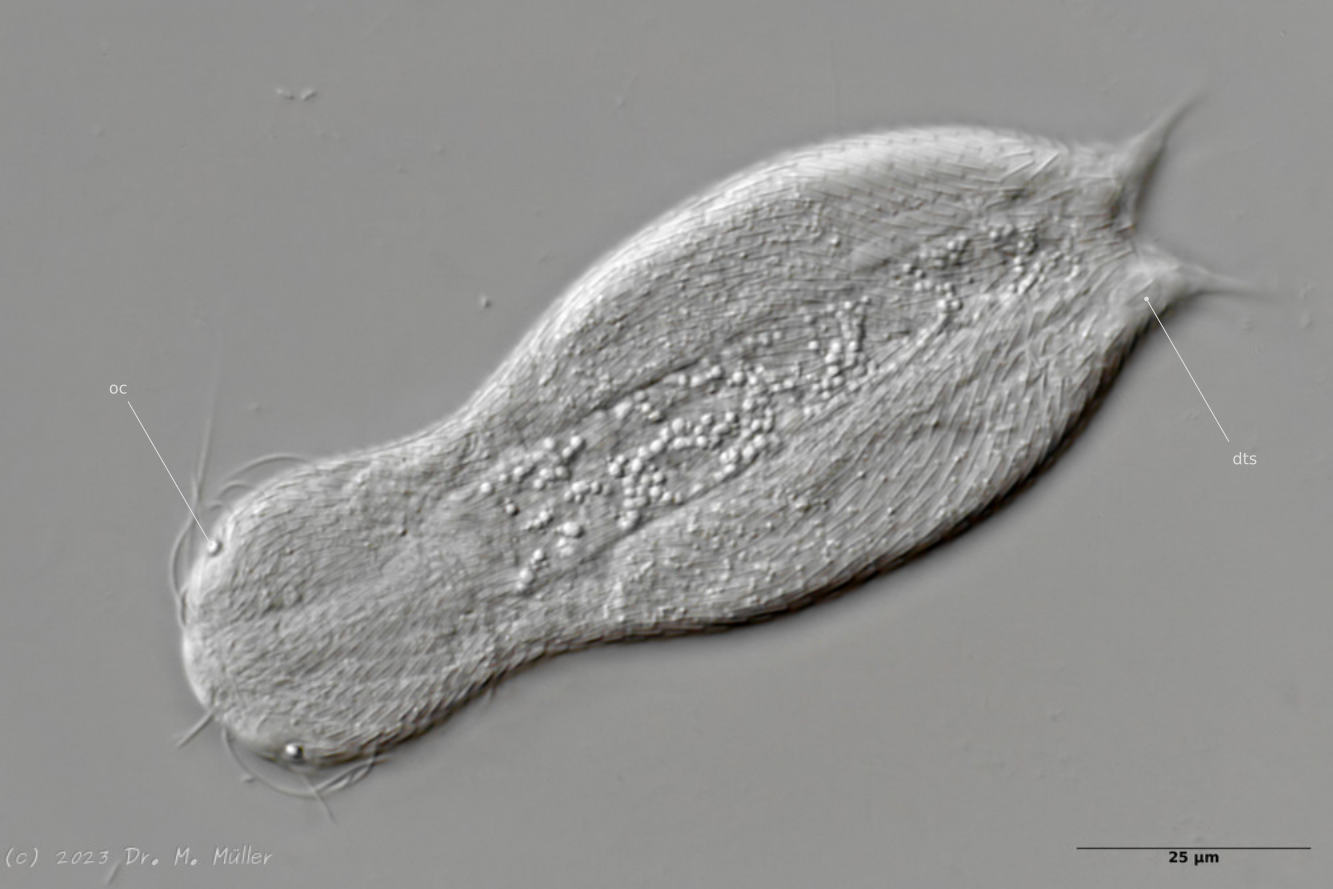
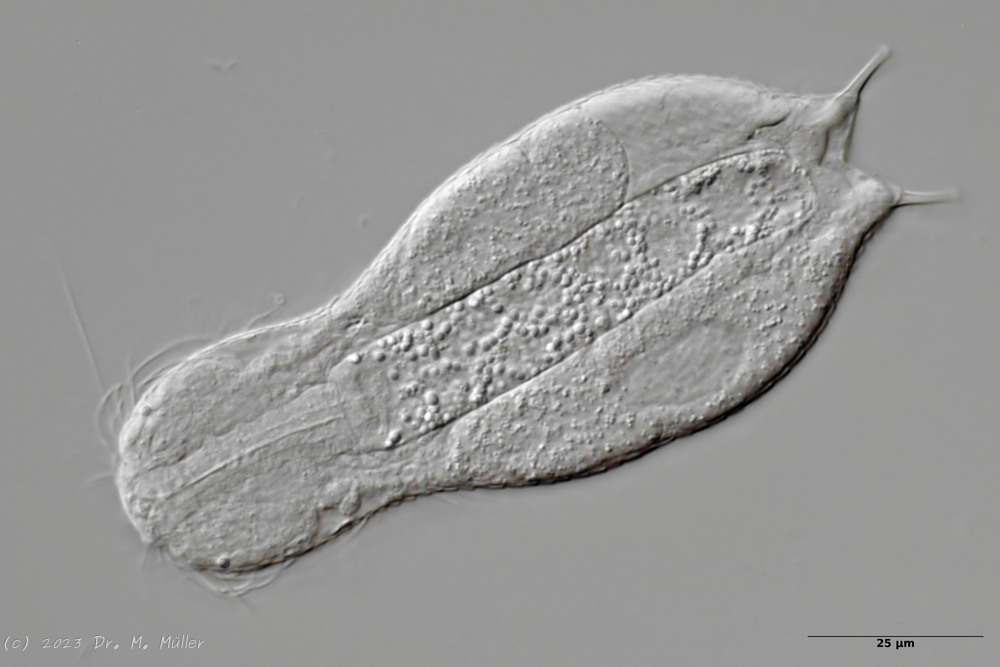
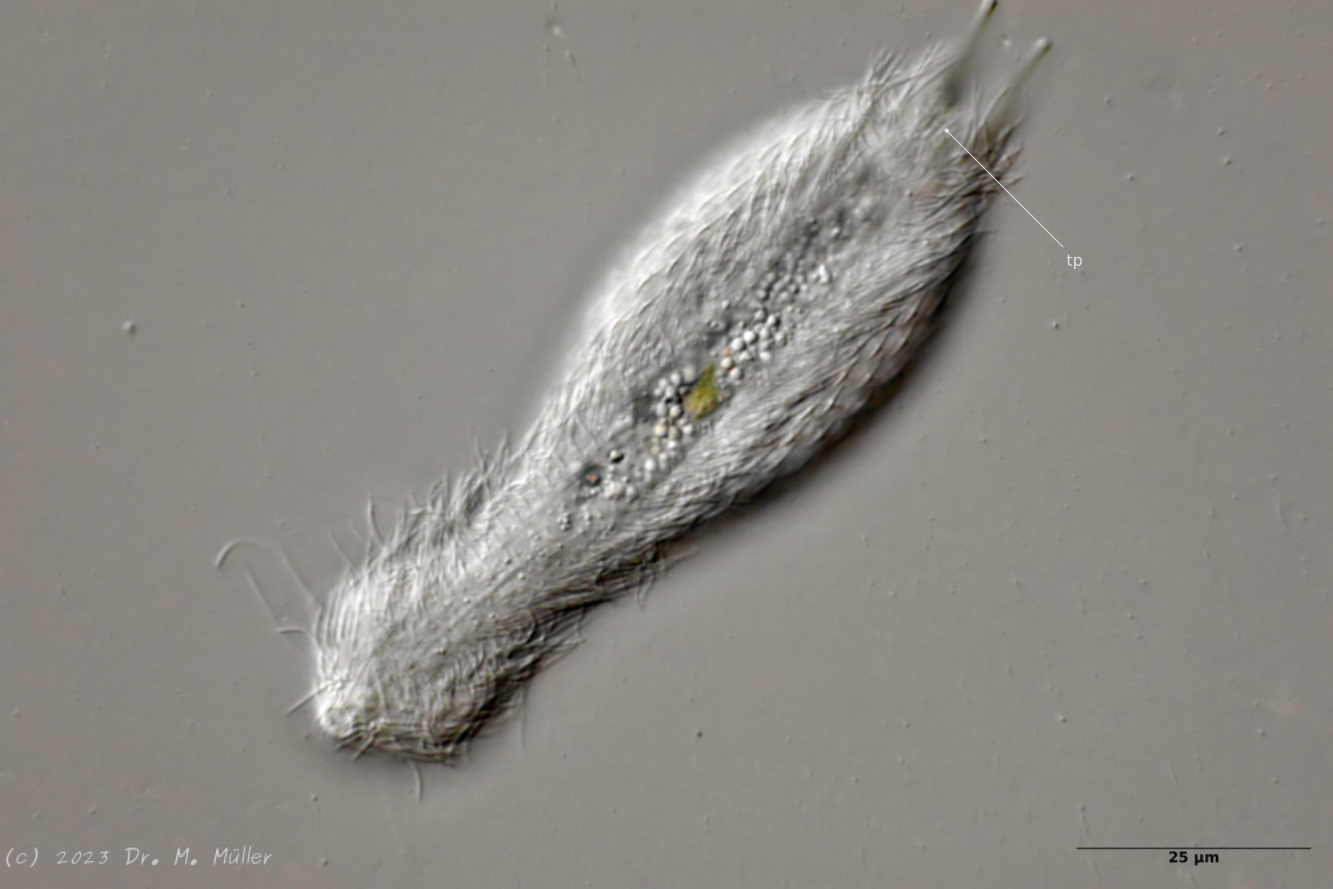
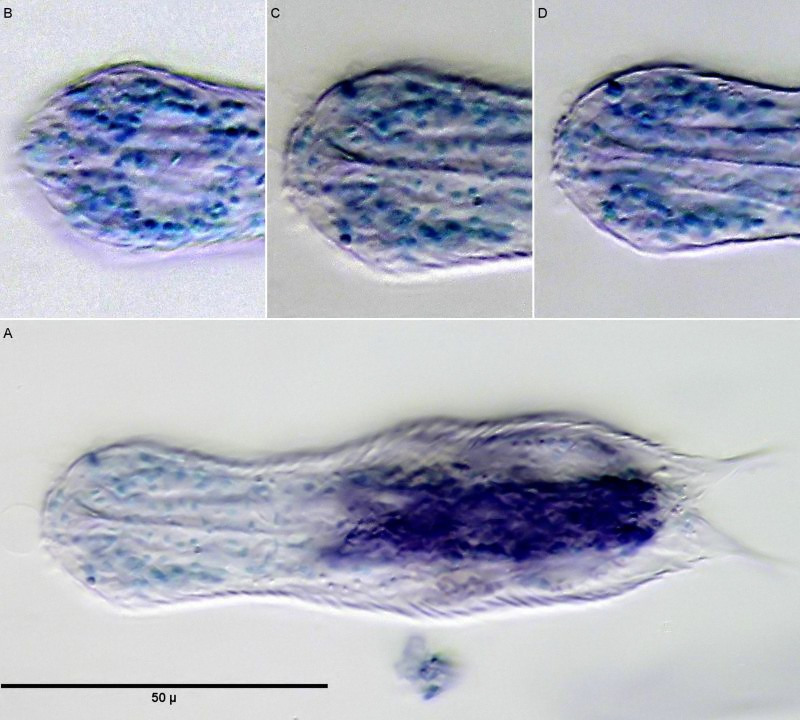
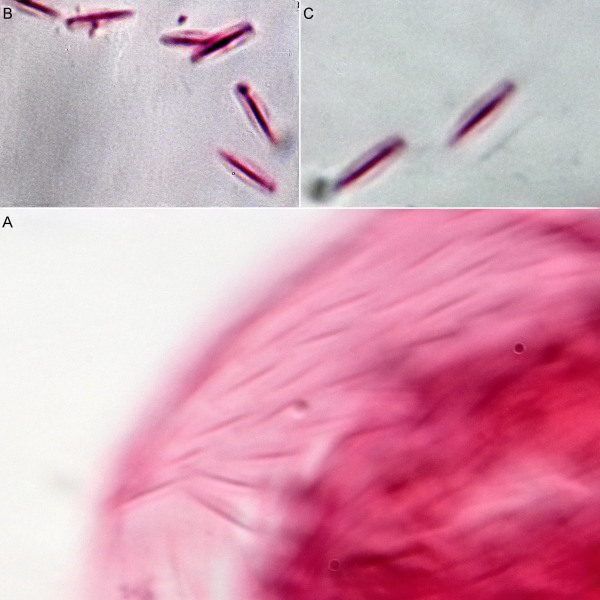
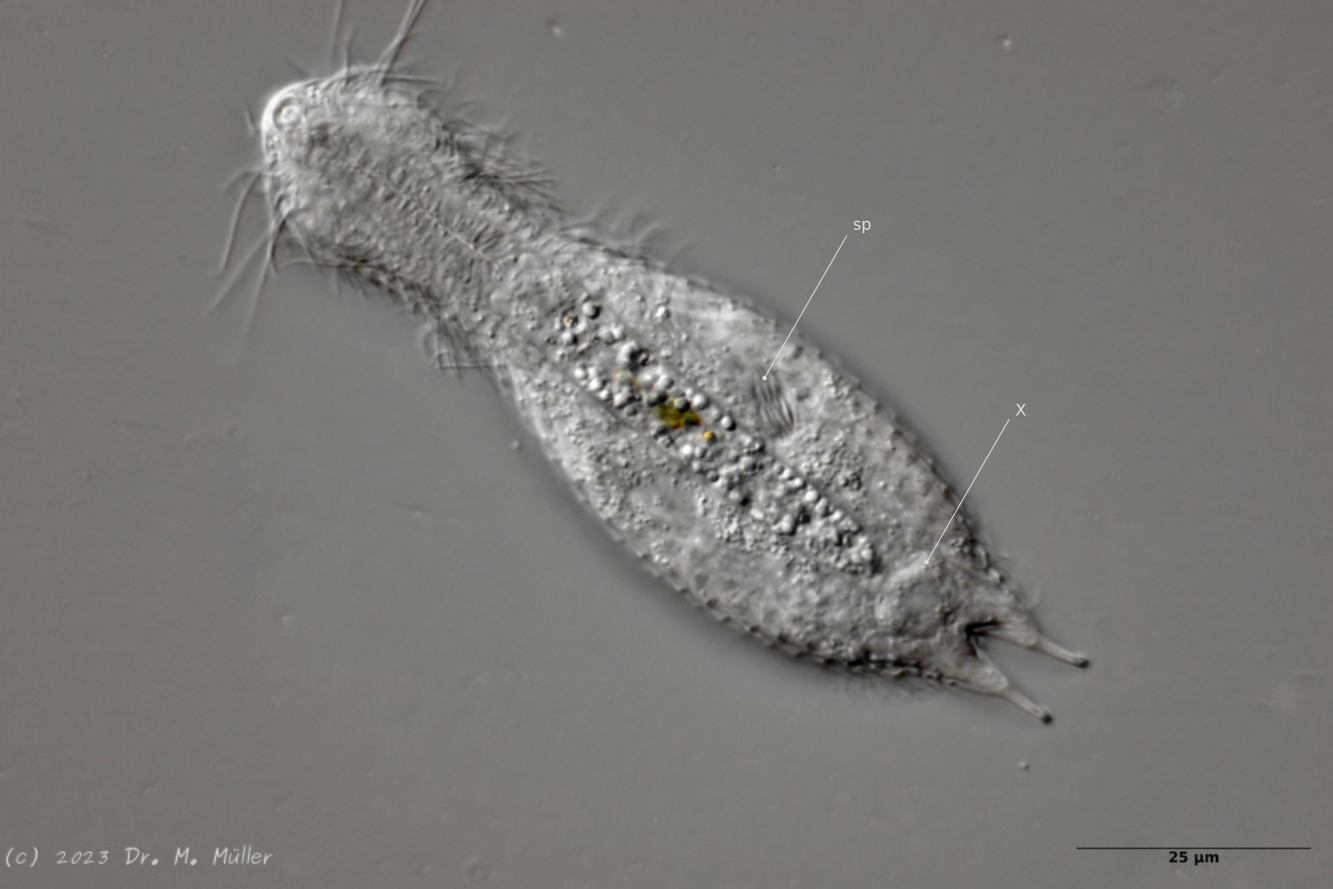
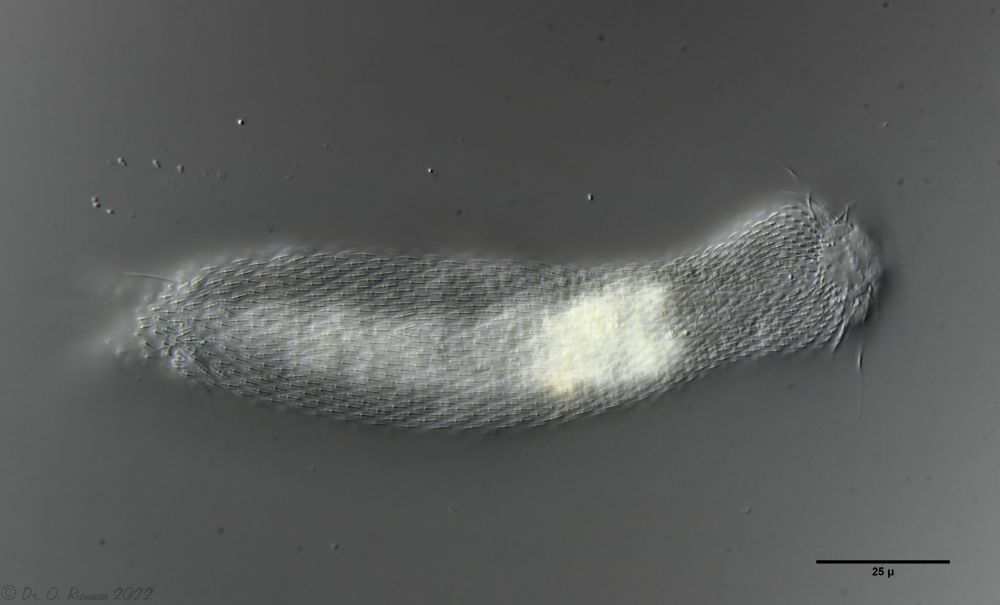
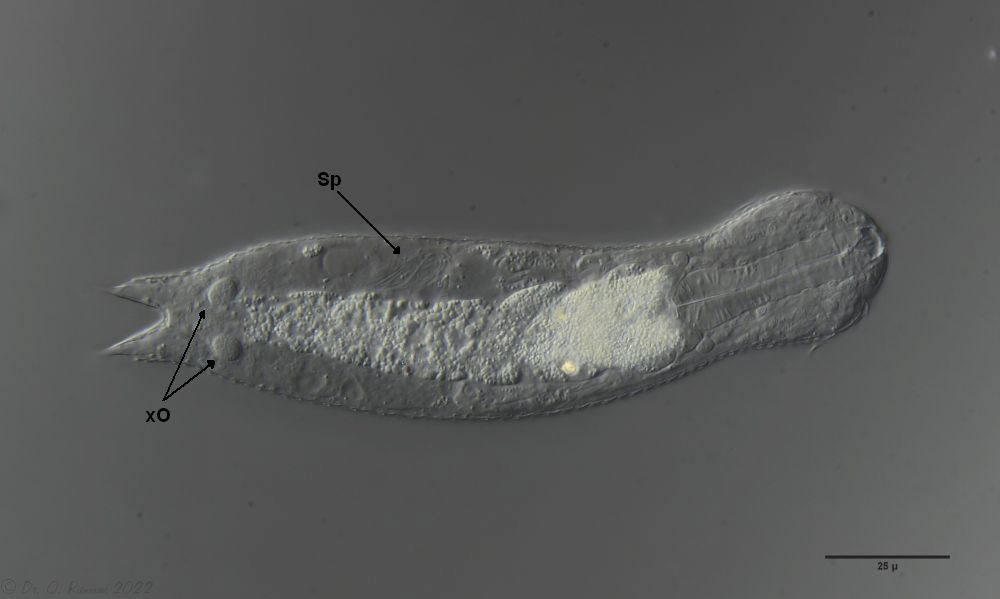
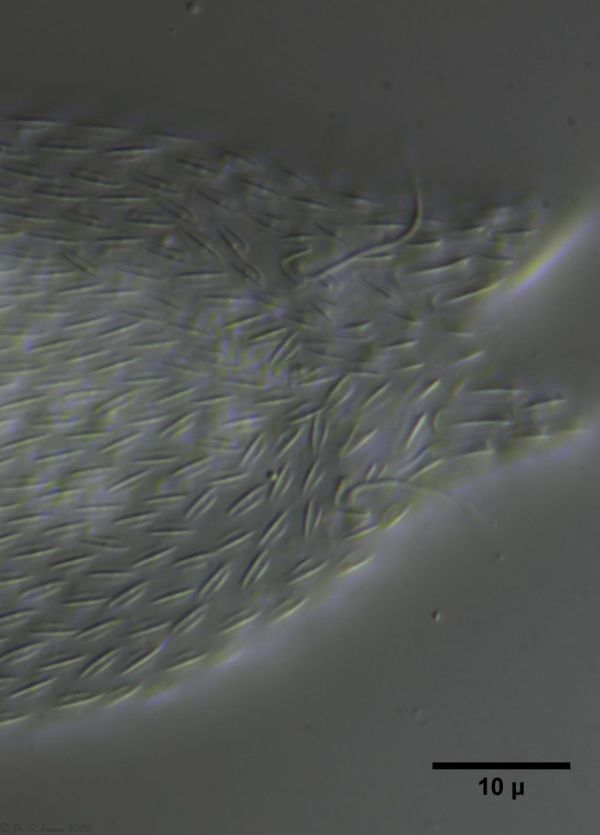
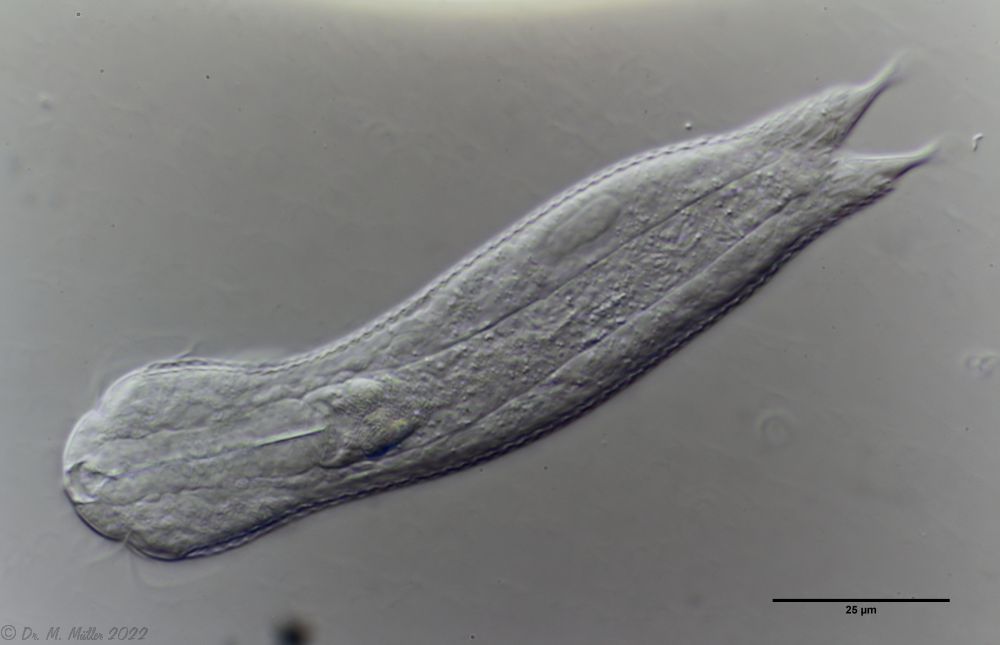
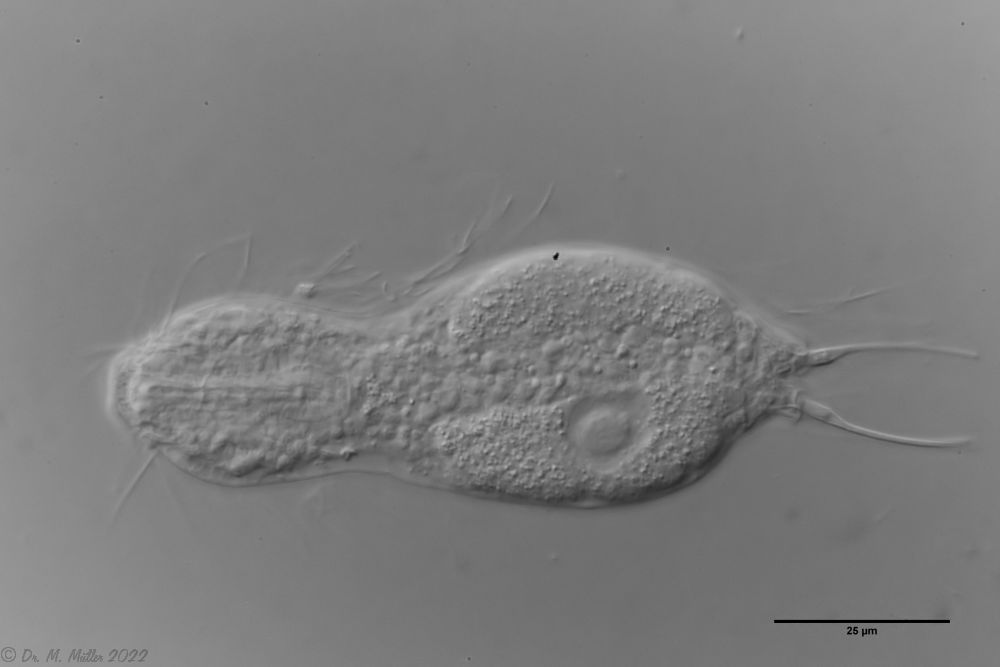
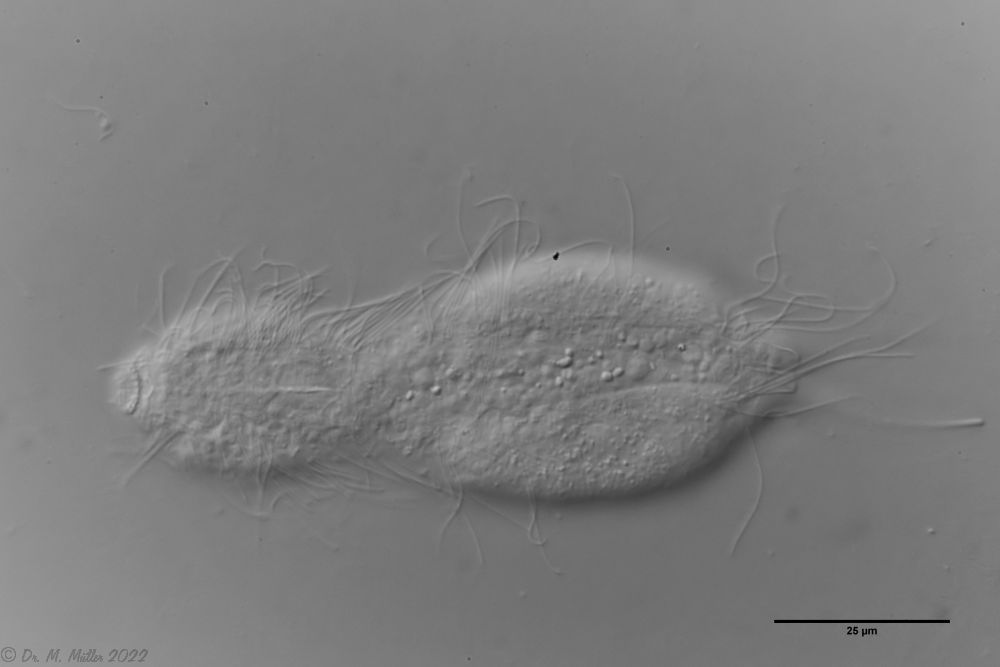
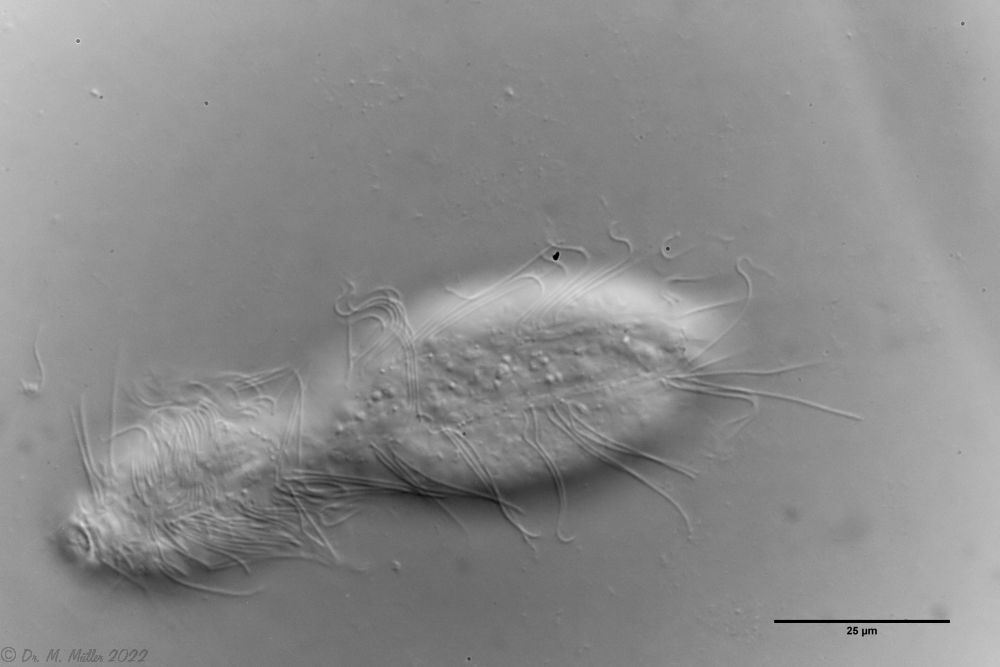
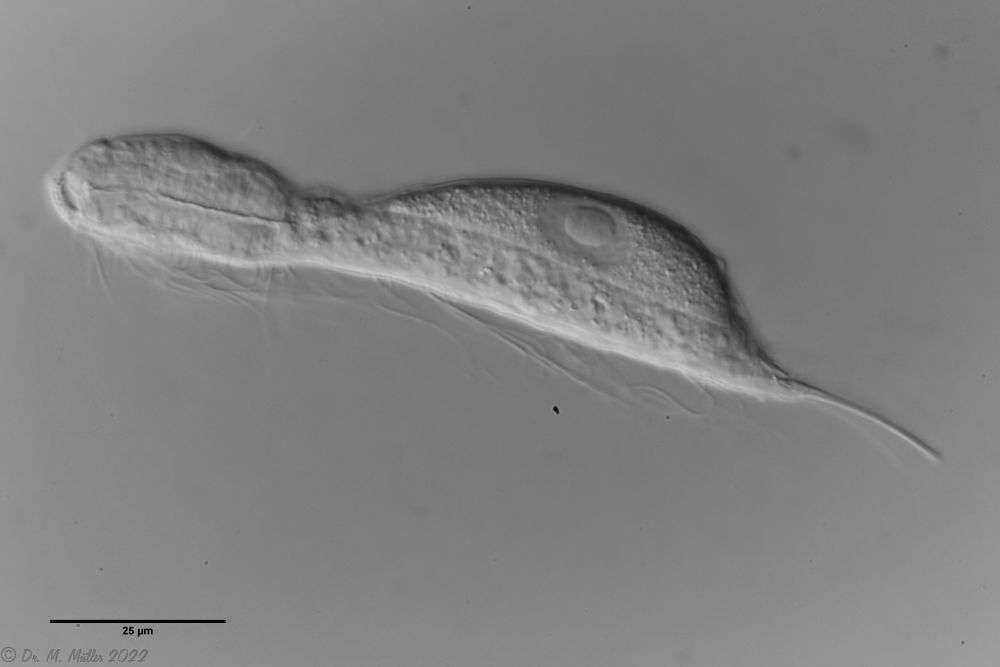
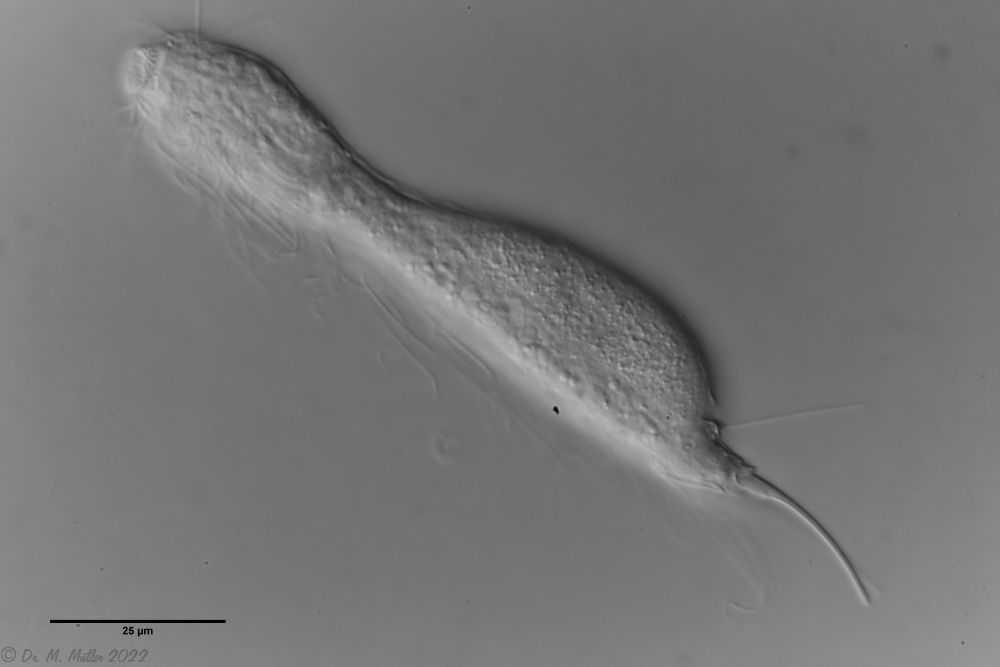
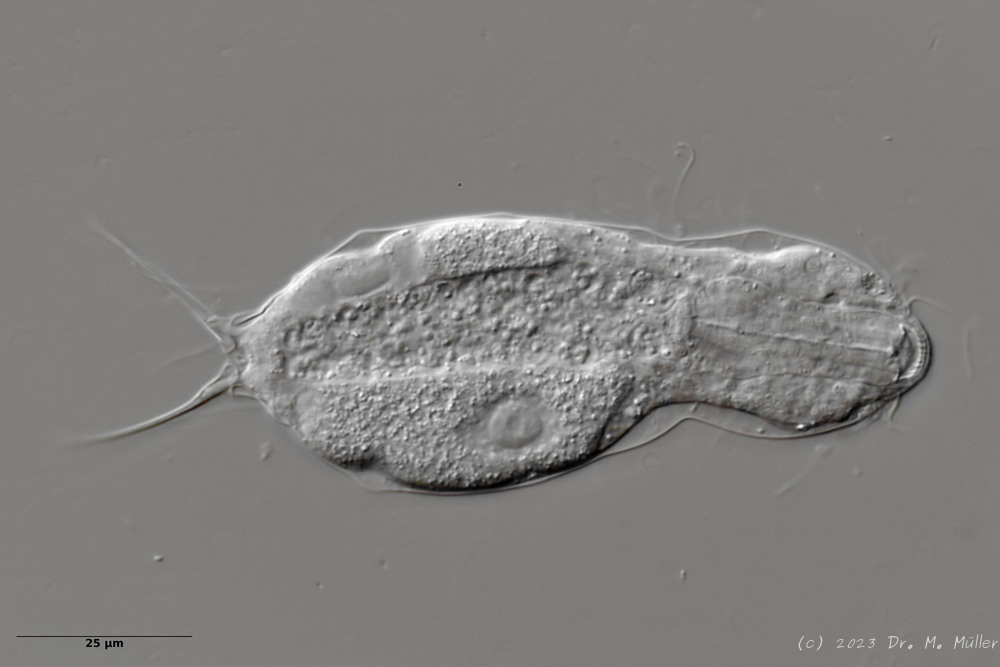
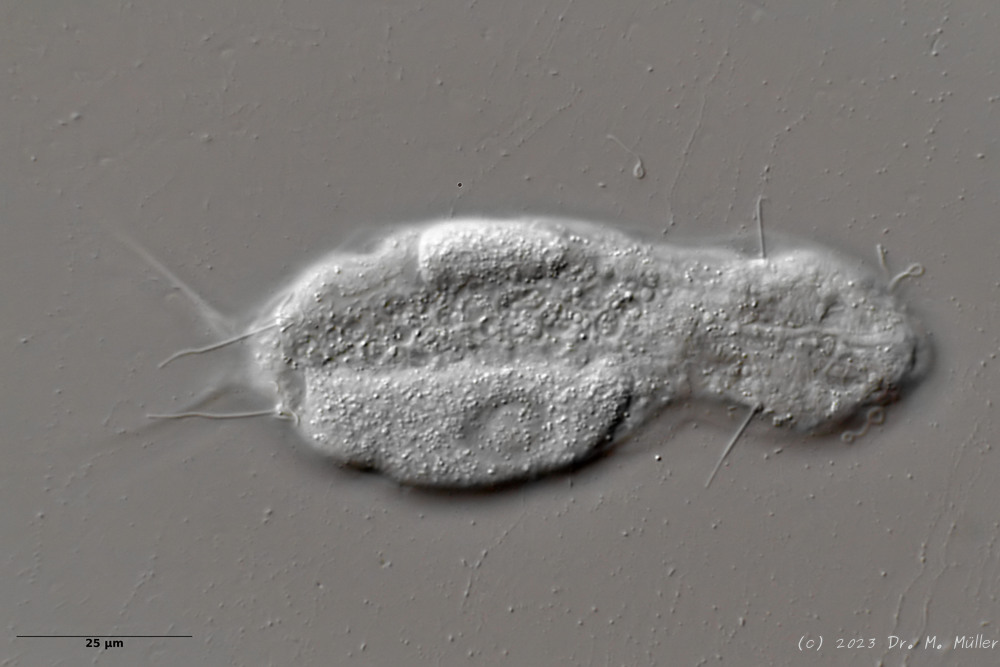
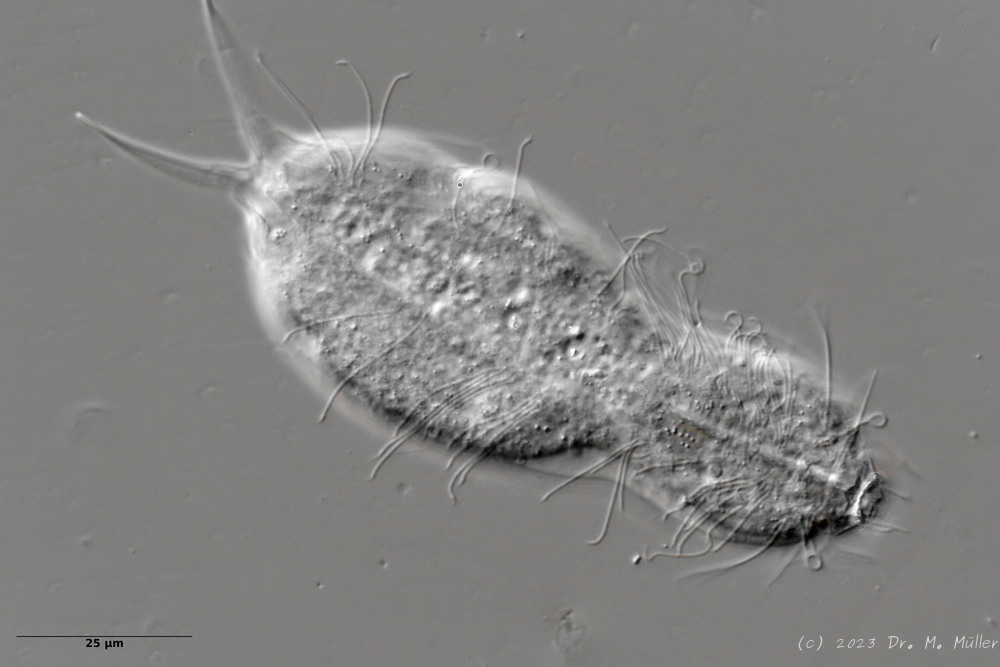
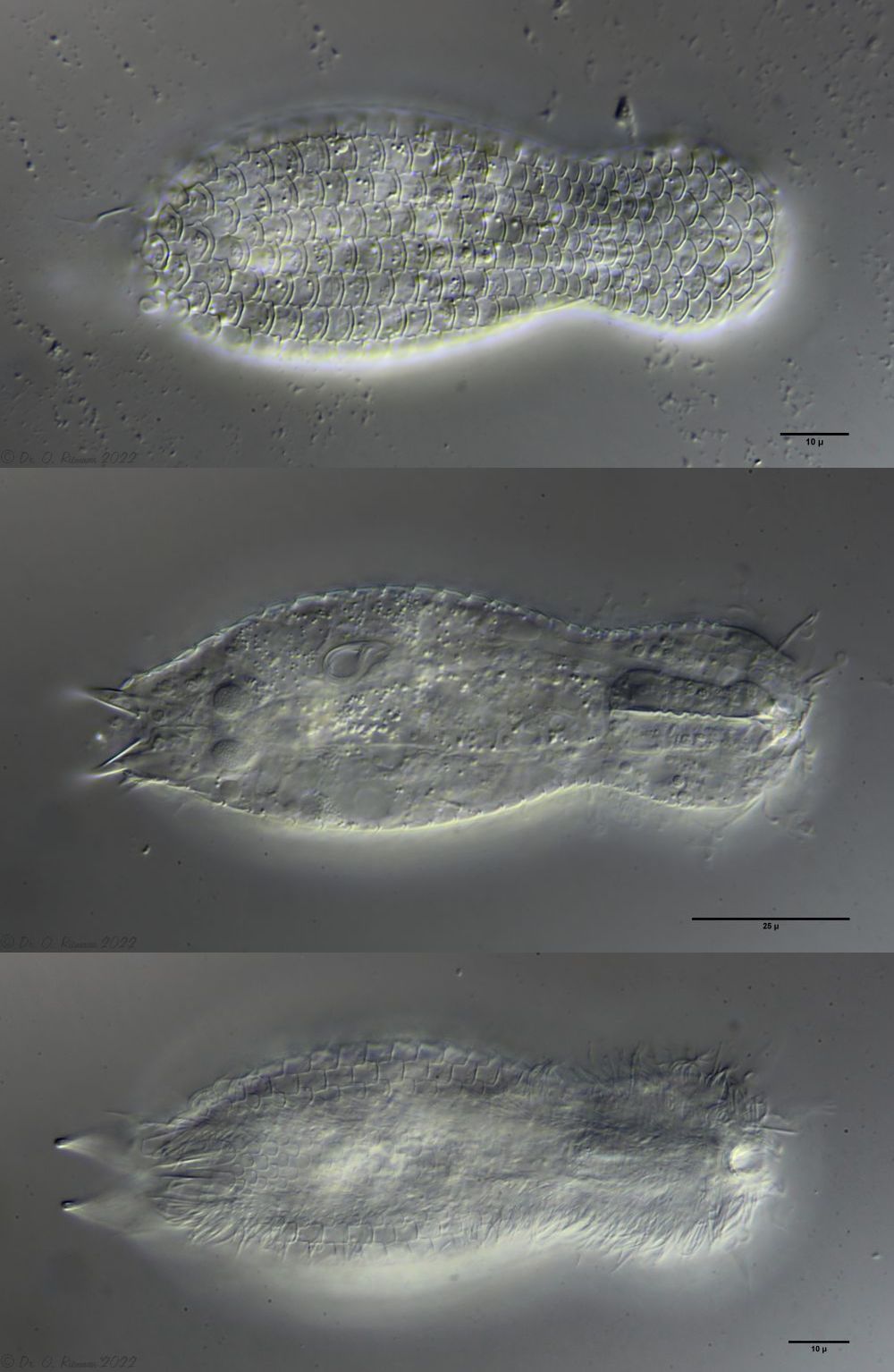 L. minor chaetifer: above dorsally with smooth scales typical of Lepidodermella;
middle in cross section with X-organ and sperm bundle;
below ventral scaling with distinct terminal plates ( Ole Riemann).
L. minor chaetifer: above dorsally with smooth scales typical of Lepidodermella;
middle in cross section with X-organ and sperm bundle;
below ventral scaling with distinct terminal plates ( Ole Riemann).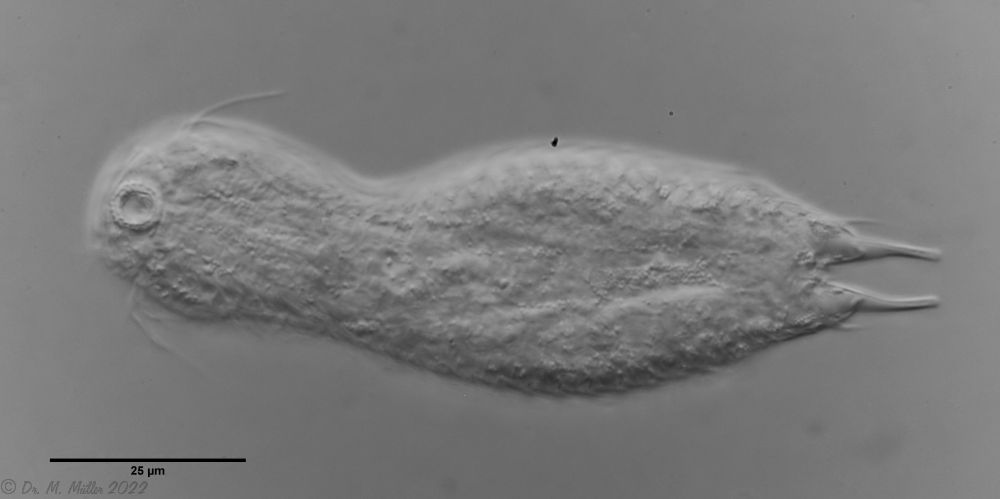 L. minor chaetifer: the pair of spines at the base of the toes is characteristic of the subspecies chaetifer
L. minor chaetifer: the pair of spines at the base of the toes is characteristic of the subspecies chaetifer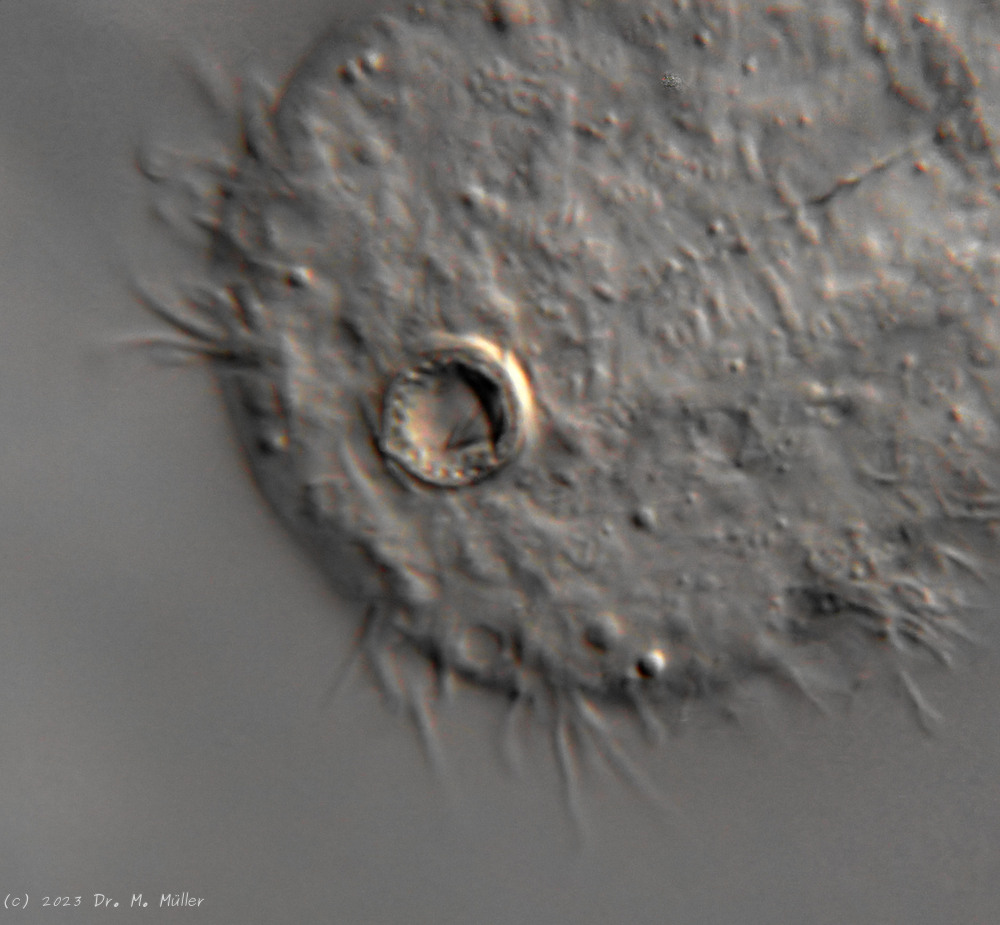
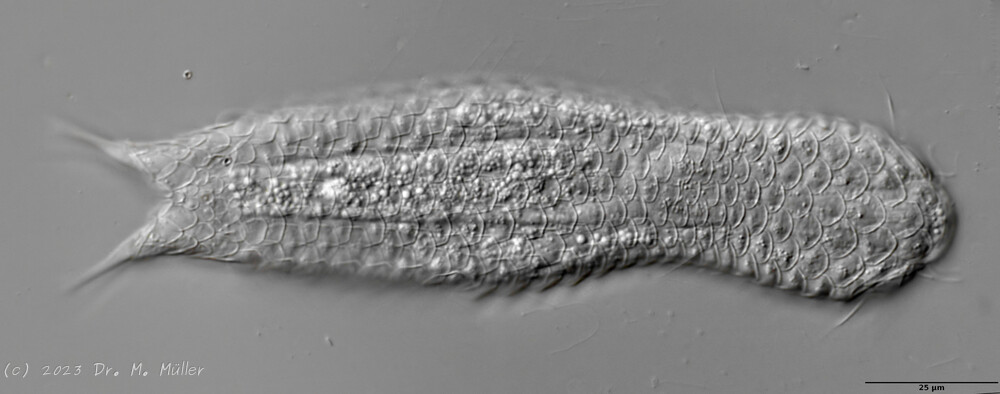 Dorsal scales
Dorsal scales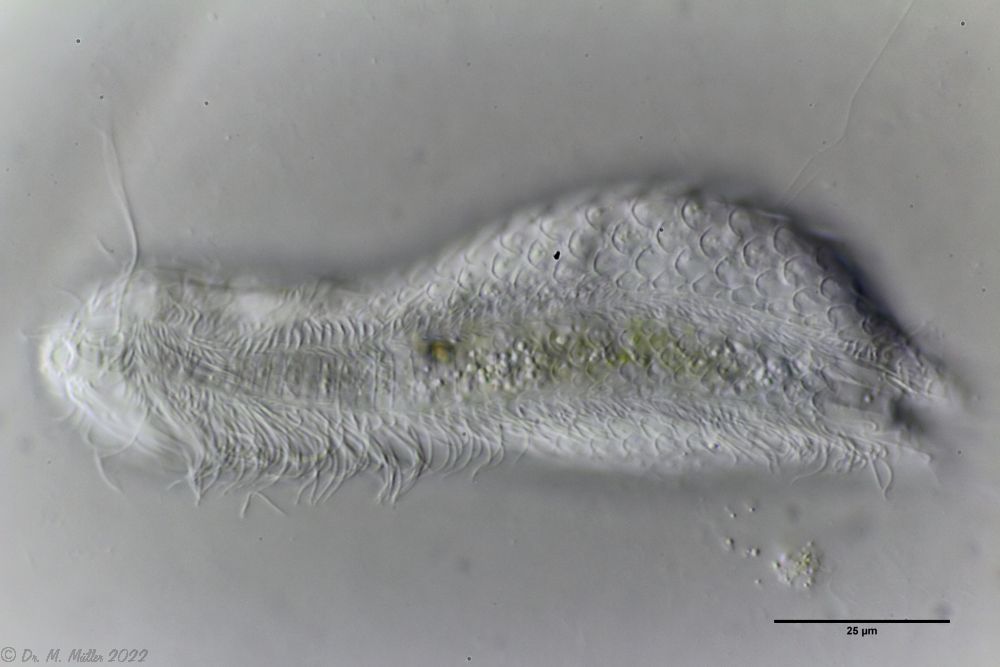 Ventral scales; large terminal plates; rectangular braces under neck
Ventral scales; large terminal plates; rectangular braces under neck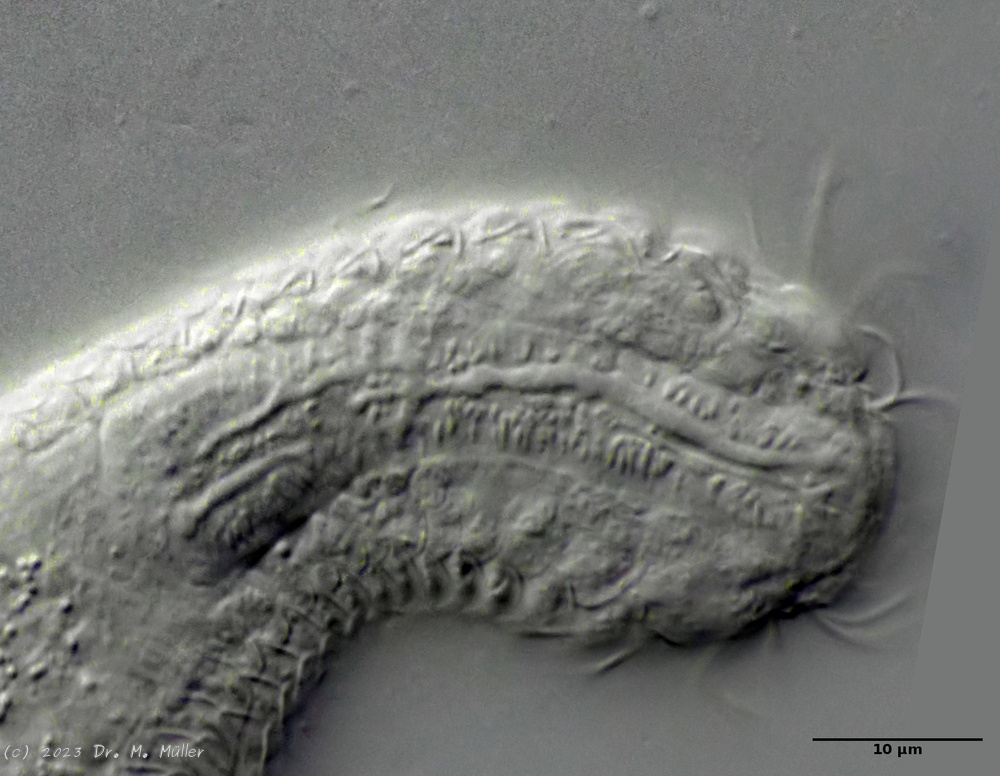
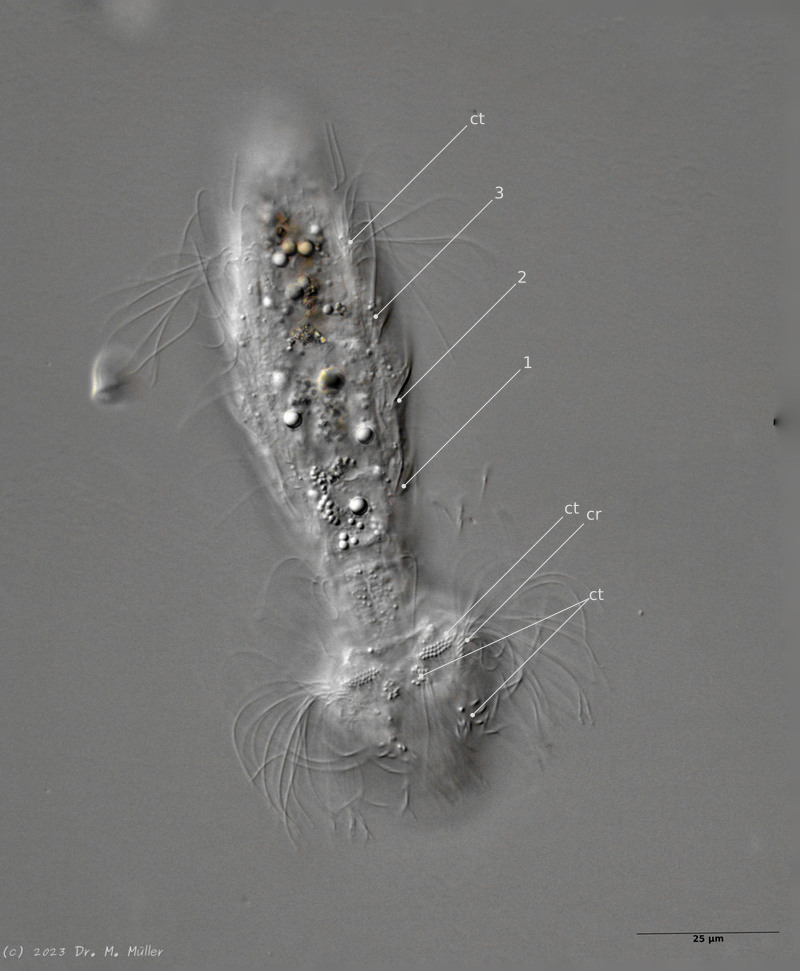
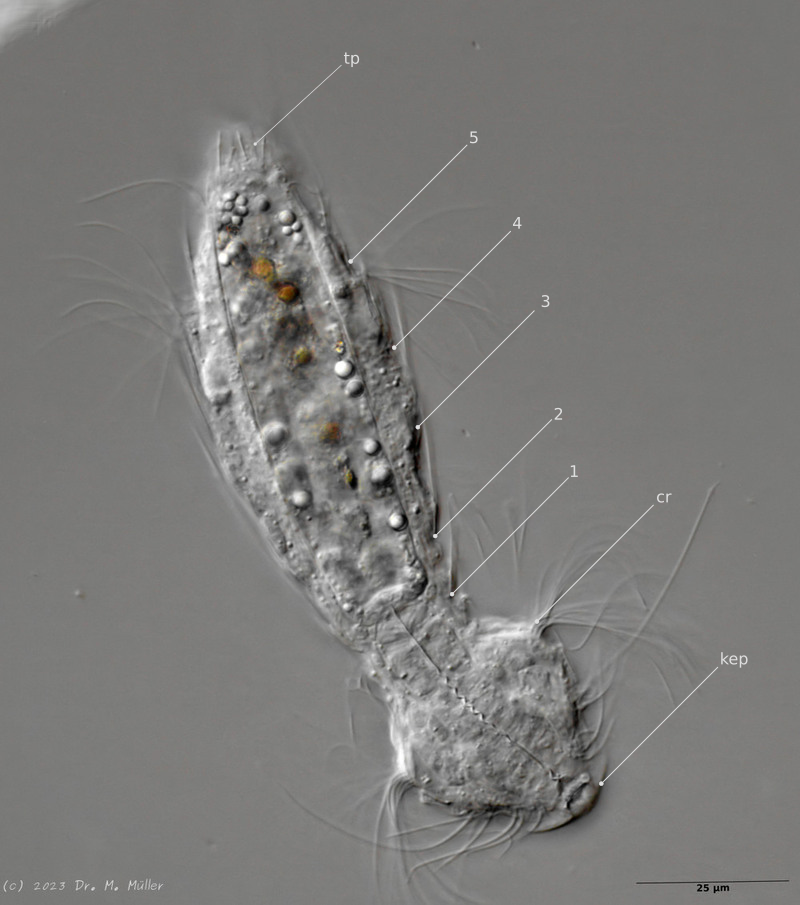
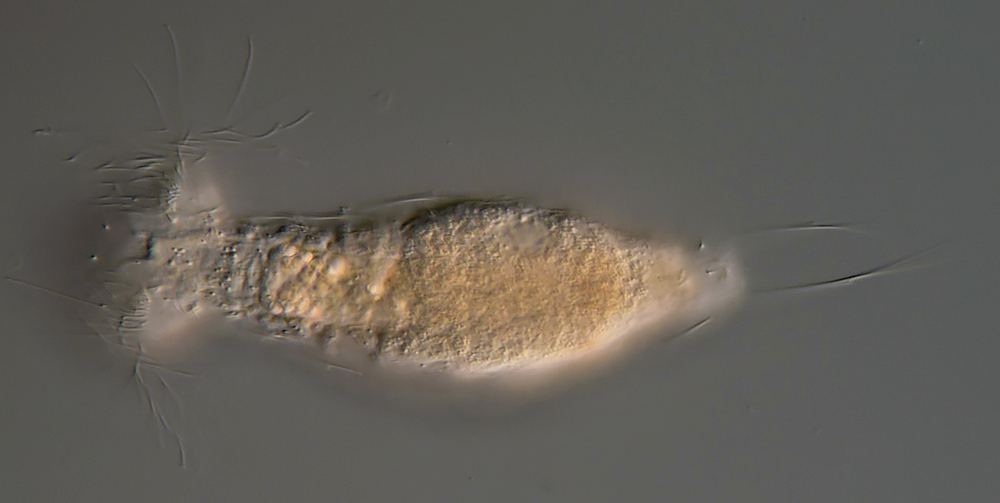
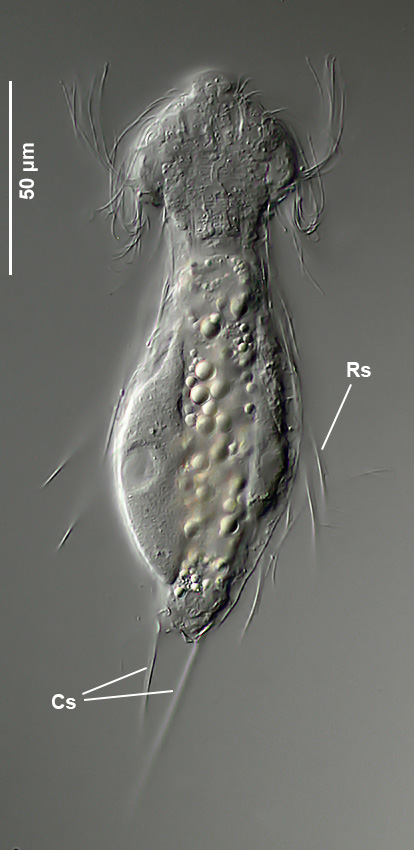


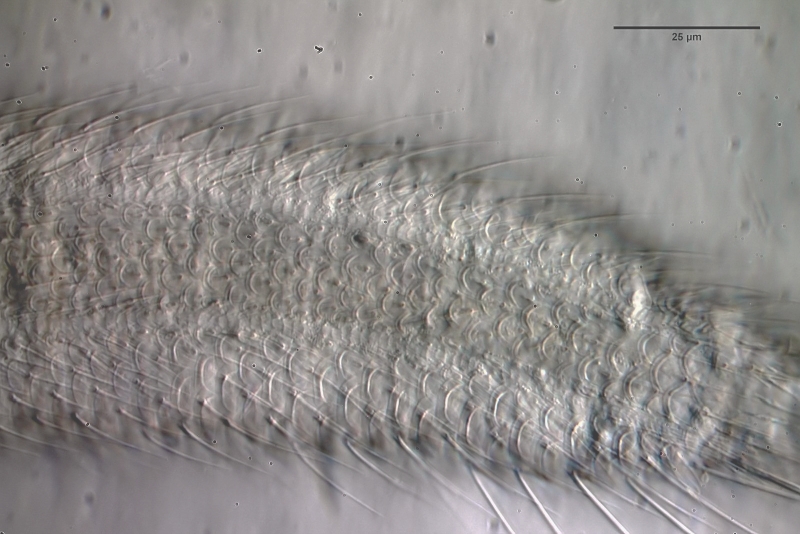
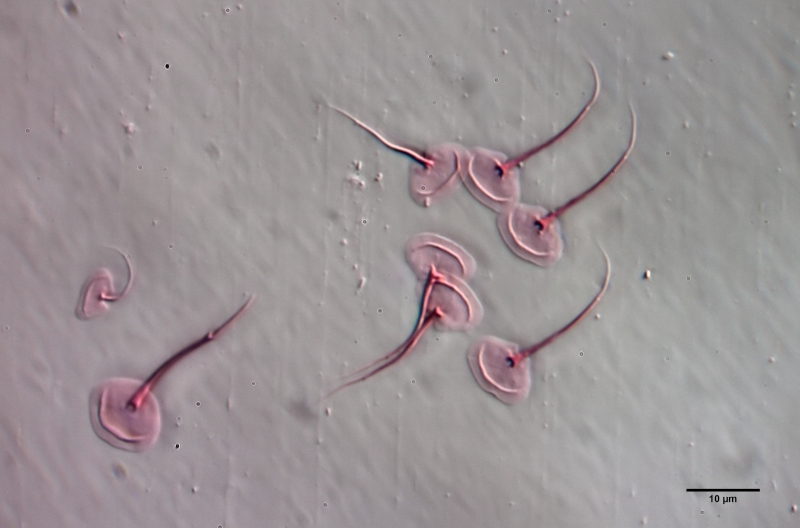
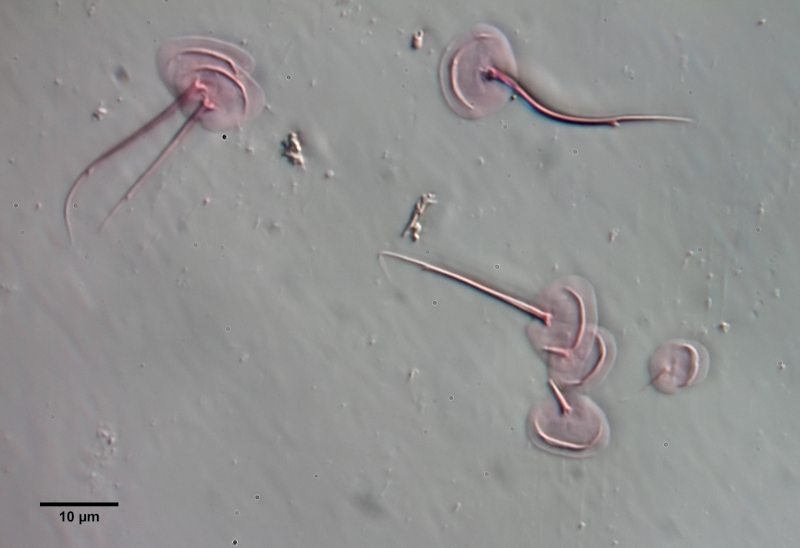
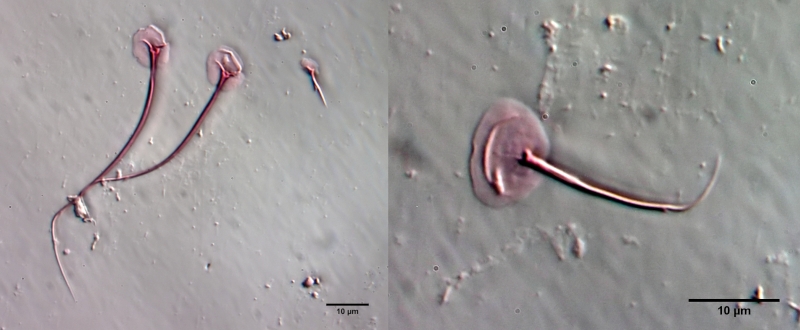
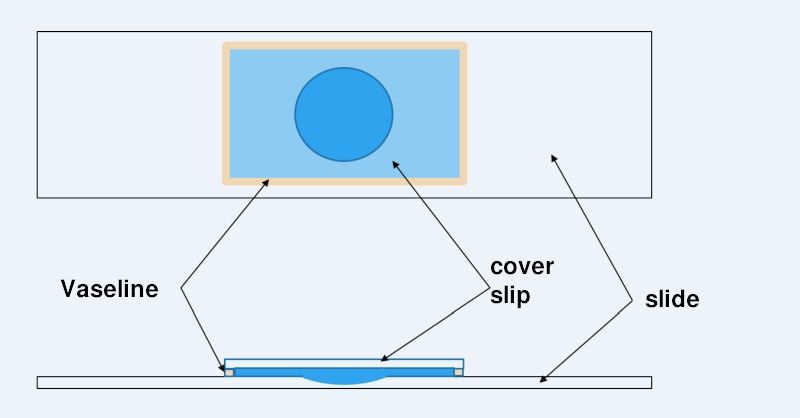




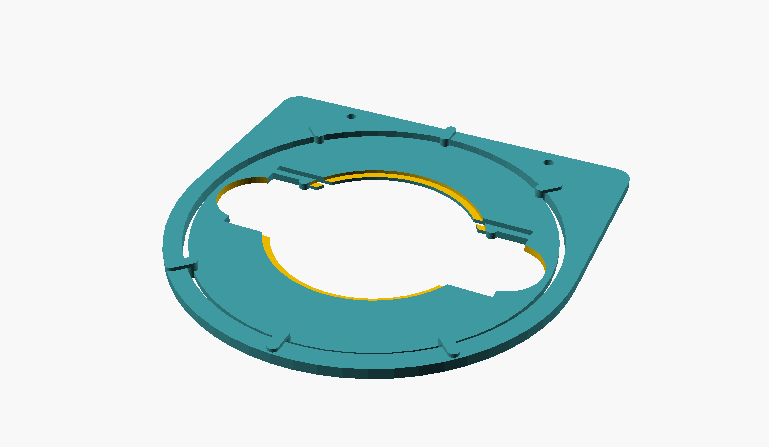



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}