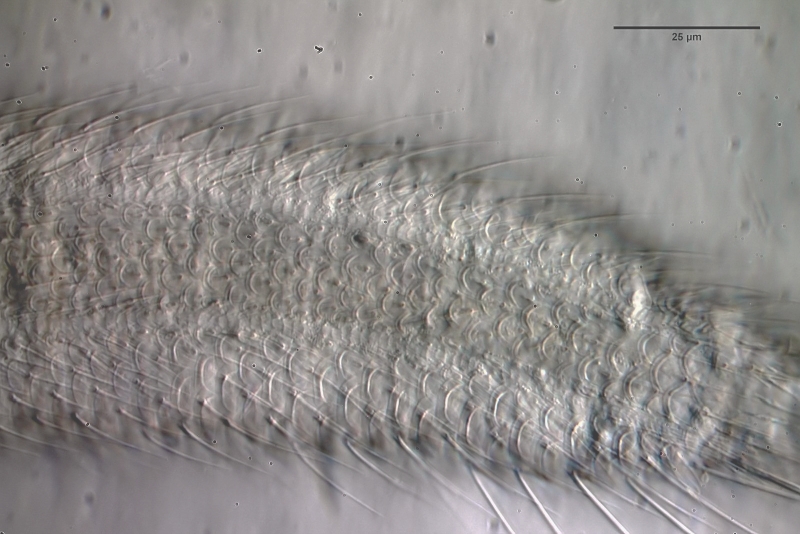
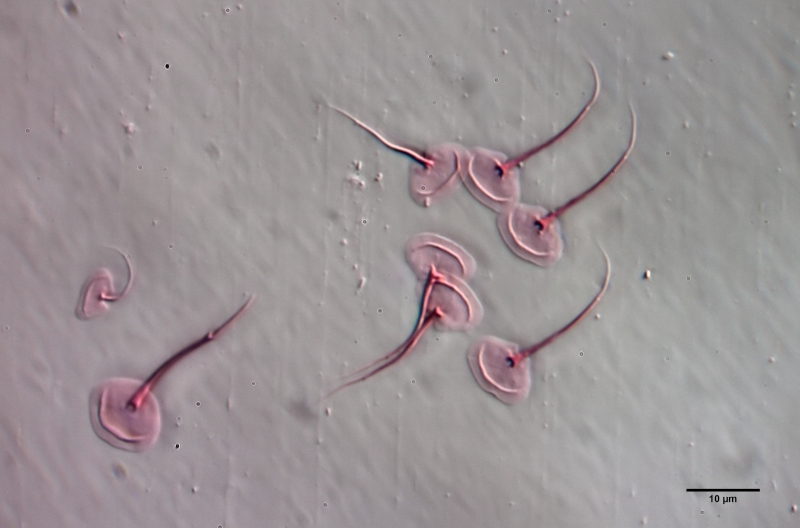
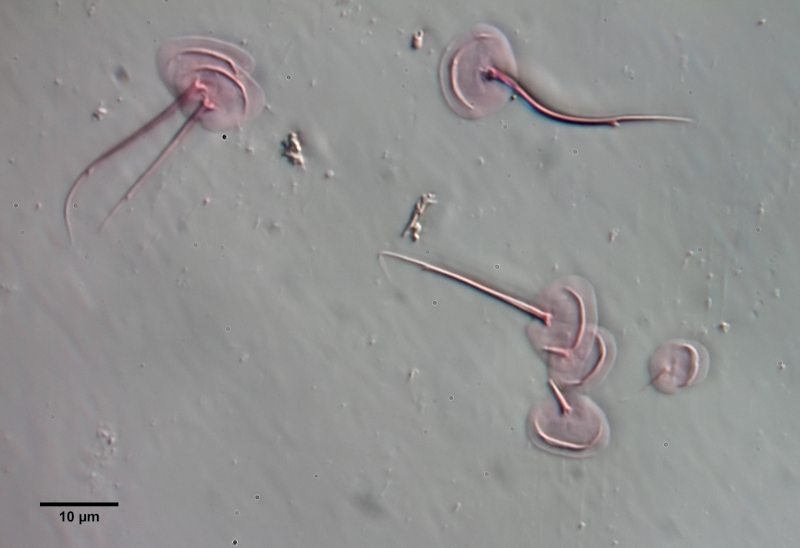
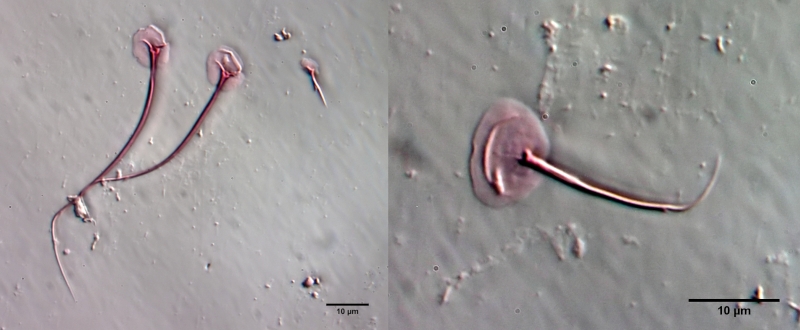
Gastrotrichen - oder Bauchhärlinge - sind mikroskopisch kleine, versteckt, in nahezu jedem Wasserkörper, lebende mehrzellige Tiere, die den meisten Menschen unbekannt sind. Dabei sind sie in vielen Habitaten die am häufigsten vertretene Mehrzeller und bilden zusammen mit den Rädertieren die Basis der mehrzelligen Fauna im Süsswasser. Dennoch ist vergleichsweise sehr wenig von Ihrer Lebensweise und ihrer Biologie bekannt und so mancher Enthusiast der mikroskopischen Untersuchung hatte selbst nach Jahren der Beschäftigung dem Leben im “Tümpel” noch keine oder nur eine sehr flüchtige Begegnung mit diesen wunderschönen und interessanten Tiere.
Deshalb möchte ich diesen scheuen Stars des Tümpels eine Web-Site widmen, in der die Süsswasser-Gastrotrichen detailliert beschrieben werden sollen. Marine Gastrotrichen werden auf diesen Seiten nicht behandelt. Deshalb ist hier - soweit nicht explizit anders angegeben - mit den Begriff “Gastrotrichen” ausschließlich die Süsswasser-Fauna gemeint.
Diese Web-Site soll sowohl dem engagierten Amateur als auch dem professionellen Beobachter bei der mikroskopischen Beobachtung und Bestimmung von Süsswasser-Gastrotrichen zur Seite stehen. Deshalb versuche ich, für möglichst alle bekannten Süsswasser-Arten Kurzbeschreibungen aus der Literatur zusammenzutragen und systematisch geordnet zu präsentieren. Nach Möglichkeit werden diese Angaben durch qualitativ hochwertige Fotos ergänzt (ggf. sind die fotografisch dokumentierte Arten mit einem “:camera:” markiert) und Identifikationshinweise für die lichtmikroskopische Artdiagnose gegeben, die sich bei meinen Beobachtungen bewährt haben.
Statistische Informationen
- Von den 397 beschriebenen Süsswasserarten beinhaltet diese Web-Site für 379 Arten Einzelseiten
- Von diesen Arten konnten bei 324 Arten keine Literaturangaben gefunden werden
- Für die übrigen 42 Arten wurde jeweils ein Steckbrief auf der Literatur übernommen
- Zu diesen Steckbriefen wurden bei 23 Arten Beobachtungsfotos hinzugefügt
Der zweite Teil beschäftigt sich mit der Biologie der Süsswasser-Gastrotrichen. Neben der Anatomie der Bauchhärlinge wird hier auch auf deren eigenartigen Lebenszyklus eingegangen.
Durch die Verwendung von Mikroaquarien war es möglich, Bauchhärlinge über eine längere Zeit (mehrere Wochen) kontinuierlich zu beobachten. Dadurch konnten einzigartige Informationen über das Verhalten und die Entwicklung von Gastrotrichen gewonnen werden.
Statistische Informationen
- Insgesamt beinhaltet diese Web-Site 4 Einzelseiten zum Thema "Biologie"
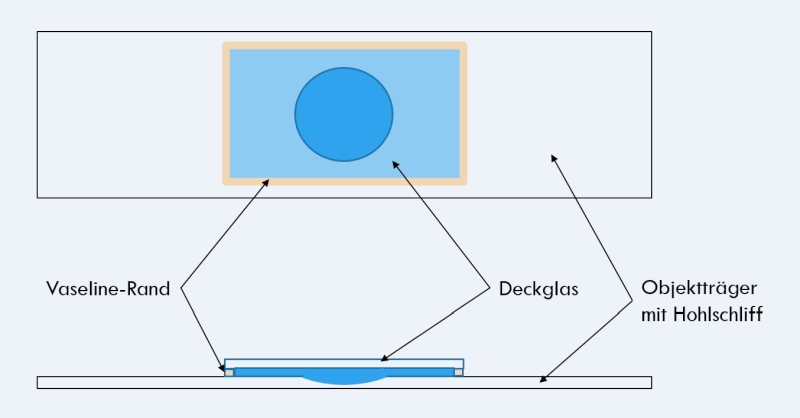
Schließlich sollen im Abschnitt “Technik” sowohl Hinweise zur nötigen bzw. von mir verwendeten apparativen Ausstattung als auch zu den von mir verwendeten Präparationstechniken gegeben werden. Hier finden sich Anleitung, die z. B. hochauflösende Beobachtungen erlauben oder Langzeitstudien der Entwicklung und des Verhaltens von Gastrotrichen durchzuführen.
Statistische Informationen
- Insgesamt beinhaltet diese Web-Site 9 Einzelseiten zum Thema "Technik"
Wie bei allen Web-Sites ist auch die vorliegende niemals fertiggestellt und konkurriert mit meinen anderen Interessen um meine knappe Zeit. Deshalb werden sich - bei aller Sorgfalt und allem Engagement - immer Lücken auftun und inhaltliche Fehler einschleichen. Die Lücken hoffe ich im Laufe der Zeit zu schließen - bei den Fehlern bin ich auf Ihre Mithilfe angewiesen. Bitte zögern Sie nicht, mich auf Probleme oder Ungenauigkeiten hinzuweisen - ich bin immer dankbar für konstruktives Feedback!
Diese Web-Site sieht sich nicht als eine wissenschaftliche Publikation. Deshalb verzichte ich zugunsten von Klarheit und Einfachheit bei manchen Themen auf die letzte “wissenschaftliche” Vorsicht. Um z. B. bei der Artbestimmung eine notwendige wissenschaftliche Sicherheit zu erreichen, wird es oft nötig sein, die Artdiagnose anhand der angegebenen Literatur zu verifizieren. Diese Web-Site soll lediglich den ersten Einstieg in das große Thema “Gastrotrichen” erleichtern.
Unterabschnitte von Die kleine Welt der Süsswasser-Gastrotrichen
gesamter Körper (incl. ventrale Zilien) mit einer weichen, kutikularen Schicht bedeckt
Schicht gut mikroskopisch von Epidermis zu unterscheiden
nur kutikulare Teile von Kopf und Haftröhrchen unabhängige Bildungen
Arten
1 Art:
Arenotus strixinoi Besonderheiten: weiche kutikulare Schicht; Zähne Dorsal: keine Schuppen, sondern geschlossene kutikulare Schicht Ventral: breites Hypostomium mit Querfurche (38-42 µm); zwei Zilienreihen; zusätzlich zei Zilien-Büschel am Kopf (median), ein Paar ZilienBüschel lateral am Kopf
Munddurchmesser ( elliptisch, zwei Zähne (9x5 µm) ): 8 x 20 µm
Dorsale Schuppen: keine Schuppen, sondern geschlossene kutikulare Schicht
Ventrale Schuppen: breites Hypostomium mit Querfurche (38-42 µm); zwei Zilienreihen; zusätzlich zei Zilien-Büschel am Kopf (median), ein Paar ZilienBüschel lateral am Kopf
Ökologie: Psammon (sandiger Grund)
Ähnliche Arten: Ähnlich zu Ichthydium, aber vollständig bedeckt mit strukturloser Kutikula (bei Ichthydium ist die Kutikula meist fein strukturiert); gut abgrenzbar durch komplexen Aufbau des Mundes
Die Gattung Aspidiophorus ist mikroskopisch sehr leicht an der scheinbaren “doppelten Kontur” des Körpers zu erkennen. Diese Doppelkontur wird durch die typischen Stilschuppen dieser Gattung erzeugt, die aus einer Basisplatte bestehen, die direkt auf der Kutikula sitzt. Aus dieser Basisplatte erhebt sich ein dünner Stiel, der an seinem Ende eine - meist größere - unbestachelte Endplatte trägt:
typische Schuppen eines Aspidiophorus, hier A. squamulosus
Daraus ergibt sich die typische doppelte Außenkontur der Tiere:
Aspidiophorus bibulbosus Dorsal: 23 Reihen zu je 40-42 kleinen Stielschuppen (2-3 µm); Stiele 0.5 µm; Form der Endplatten unbeschrieben Ventral: 8-9 Reihen kleiner Stielschuppen; keine Terminalplatten; Halsregion nackt
Aspidiophorus heterodermus levantinensis Dorsal: rhombische Stielschuppen (3µm lang), 41-42 pro Reihe, letzte Reihe in der Mitte mit Stacheln (9-14µm), lateral an Zehenbasis ein Paar langer Stacheln (16-18µm), manchmal mit einem weiteren Paar Stacheln oder verlängerten Schuppen Ventral: vZwf. mit einem Paar terminaler Stacheln; sehr viele winzige Kiele
Aspidiophorus longichaetus Dorsal: 13-14 Reihen mit je 24 Stielschuppen; lange, ungekielte, rhombische Endplatten (6-7µm); hintere 4 Endplatten 10µm, stachelartig zugespitzt; Stiellänge 1.2µm; Afterregion: 3 einfache, schuppenlose Stacheln (18-20µm), Zehenausschnitt überragend; Auf der Zehenbasis langes Stachelpaar (39µm) Ventral: vZwf nackt, 2 terminale, schuppenlose Stacheln (10µm)
Aspidiophorus microlepidotus Dorsal: 50 Reihen mit je 120 winzigen Stielschuppen; am Hinterende 4 kurze Stacheln Ventral:
Aspidiophorus microsquamatus Dorsal: 40-45 Reihen mit je 40-45 winzige Stielschuppen; Hinterende nackt; Ventral: zwei große Terminalplatten, wenige Reihen Stielschuppen, sonst nackt
Aspidiophorus oculifer Dorsal: 17-22 Reihen mit je 36-41 kleinen Stielschuppen; Endplatten 2.5-4 µm; Stiele 0.6-1 µm; keine Stacheln Ventral: 2 gekielte und bestachelte Terminalplatten (6.5-10.5µm); 5-12 Reihen 1,5-3µm langer Kiele
Aspidiophorus ontarionensis Dorsal: 9 Reihen mit je 16-17 Stielschuppen; Endplatten (6-7 x 2-3 µm) vorne gerundet, hinten eingeschnitten, keine Kiele; letzten 2-3 Reihen laufen in 20 µm Stacheln aus; lateral an Zehen ein Paar Stacheln (25 µm) Ventral: vZwf mit 25 Reihen winzigen Stielschuppen
Aspidiophorus ophiodermus Dorsal: 41-44 Reihen zu je 48-50 kleinen Stielschuppen; Endplatten elliptisch, zugespitzt mit Kiel, überlappend, 3 x 1.5 µm; zwei 5x2 µm große Stielschuppen an der Furka-Innenseite Ventral: vZwf mit zwei Terminalplatten (7x4 µm) und 10 Reihen schmaler, distal zugespitzter Kielschuppen (4x0.75µm)
Aspidiophorus paradoxus Dorsal: 25 Reihen mit je 40-45 rhombischen Stielschuppen mit seitlichen und medianen Kielen (4x6 µm); Stiele 6-7 µm; Endplatten der letzten Endreihen oft vergrößert (12-17 µm); Hinterende frei; zwischen Zehen 4 kurze Stacheln auf ovalen Schüppchen Ventral: vZwf mit Stielschuppen bedeckt
Aspidiophorus pleustonicus Dorsal: 45-53 Reihen zu je 29-32 Stielschuppen (5-7 µm); kein Kiel, nicht rhombisch, ähnliche Form wie bei
P. rhomboides
; Zehenbasis dorsal mit Kielen Ventral: 6 gekielte Terminalplatten; am Hinterende 10-12 Reihen von Kielen (4-4.5 µm), sonst nackt
Aspidiophorus polonicus Dorsal: 18-20 Reihen mit je 42-45 kleinen Stielschuppen; Stiele 1.5 µm; Endplatten 2-3.5 µm lang, proximal verstärkter Rand, distal zugespitzt, schwacher Mittelkiel; keine Stacheln Ventral: vZwf. nackt, bis auf wenige längliche Stielschuppen; zwei schmale Terminalplatten(6.5-11.5 µm)
Aspidiophorus pori Dorsal: 53 Stielschuppen (3 µm) pro Reihe; 3 Paar Stacheln an der äußeren Zehenbasis (20-22 µm) Ventral: vZwf sehr viele Stielschuppen; ein Paar terminaler Stacheln (8-10 µm) und viele kürzere Stacheln, die in den Zehenausschnitt ragen
Aspidiophorus schlitzensis Dorsal: 25-30 Reihen mit je 60-80 winzigen Stielschuppen; Endplatten ungekielt (0.5-2 µm), länglich, distal zugespitzt; Hinterende mit verlängerten Kielschuppen Ventral: vZwf. zahlreiche Längsreihen winziger Stielschuppen; zwei gekielte Terminalplatten (6-7 µm)
Aspidiophorus slovinensis Dorsal: 15-17 Reihen mit je 40-45 kleinen Stielschuppen; Endplatten 2-3 µm;, abgerundet; keine Stacheln Ventral: vZwf. mit Stielschuppen; großes trapezförmiges Hypostomium mit Querfurche (14µm lang)
Aspidiophorus squamulosus Dorsal: 12-15 Reihen mit je 40-42 Stielschuppen mit rhombischen Endplatten ohne Mittelkiel (3-4 x 5-6 µm); Schuppenpanzer endet in Afterregion; Hinterende und Zehen nackt; 3 Paar kurze Stacheln an Zehenbasis (8-10 µm) Ventral: 2 terminale Kiele (5-6 µm), 12 Reihen Kiele ohne Basisplatten
Aspidiophorus tatraensis Dorsal: 15-20 Reihen mit je 38-40 sehr kleinen Stielschuppen; Endplatten sehr zart und ungekielt (2.5-3 µm); stark gebogene Stiele -> Schuppen stehen borstenartig ab Ventral: vZwf. 9-10 Reihen Kiele; vorne meist nackt; keine Terminalplatten
Aspidiophorus tetrachaetus Dorsal: 15-17 Reihen mit je 33-40 Stielschuppen; Endplatten ungekielt 2-5µm, distal und proximal abgerundet; Zehenbasis nackt; 2 Paar dicke Stacheln (8-10µm und 4-5µm) auf ovalen Schuppen mit zwei Kielen Ventral: 2 terminal Kiele (7.5-11µm); in Darmregion 8-9 Reihen winziger Kiele, sonst nackt
Unterabschnitte von Aspidiophorus
Aspidiophorus aster
Taxonomie
Ordnung: CHAETONOTIDA Remane, 1925
UnterOrdnung: PAUCITUBULATINA d'Hondt, 1971
Familie: CHAETONOTIDAE Gosse, 1864
Unterfamilie: CHAETONOTINAE Kisielewski, 1991
Gattung: Aspidiophorus Voigt, 1903
Art: aster
Länge ( flaschenförmig ):
Breite:
Kopfbreite ( fünflappig ):
µm
Furkalänge:
Haftröhrchen:
Pharyx ( zylindrisch ):
Munddurchmesser ( rund ): unbekannt
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Martin,L.V. 1981. Gastrotrichs found in Surrey. Microscopy 34: 286-300.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Grosso,L.E. 1973. Notas sobre Gastrotrichos Argentinos II. Neotropica 19(59): 87-89.
Grosso,L.E. 1973. Notas sobre Gastrotrichos Argentinos I. Chaetonotus (Zonochaeta) guruguetoi sp. nov. y Aspidiophorus brahmsi sp. nov. Physis 32B: 133-137.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Dorsale Schuppen: rhombische Stielschuppen (3µm lang), 41-42 pro Reihe, letzte Reihe in der Mitte mit Stacheln (9-14µm), lateral an Zehenbasis ein Paar langer Stacheln (16-18µm), manchmal mit einem weiteren Paar Stacheln oder verlängerten Schuppen
Ventrale Schuppen: vZwf. mit einem Paar terminaler Stacheln; sehr viele winzige Kiele
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Grosso,L.E. and Drahg,F. 1983. Gastrotricos dulceacuicolas de la provincia de Tucman. I. Chaetonotus soberanus sp. nov. y Aspidiophorus lilloensis sp. nov. Neotropica 29: 189-193.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Pharyx ( zylindrisch, mit kleinen terminalen Schwellungen ): 30 µm - 35 µm
Munddurchmesser ( rund ): 3 µm
Dorsale Schuppen: 9 Reihen mit je 16-17 Stielschuppen; Endplatten (6-7 x 2-3 µm) vorne gerundet, hinten eingeschnitten, keine Kiele; letzten 2-3 Reihen laufen in 20 µm Stacheln aus; lateral an Zehen ein Paar Stacheln (25 µm)
Ventrale Schuppen: vZwf mit 25 Reihen winzigen Stielschuppen
Ökologie: Schlammbewohner
Ähnliche Arten: A. longichaetus
: Kopf eher dreilappig, keine medianen Stacheln
Dorsale Schuppen: 41-44 Reihen zu je 48-50 kleinen Stielschuppen; Endplatten elliptisch, zugespitzt mit Kiel, überlappend, 3 x 1.5 µm; zwei 5x2 µm große Stielschuppen an der Furka-Innenseite
Ventrale Schuppen: vZwf mit zwei Terminalplatten (7x4 µm) und 10 Reihen schmaler, distal zugespitzter Kielschuppen (4x0.75µm)
Pharyx ( zylindrisch, an Enden etwas geschwollen ): 76 µm - 85 µm
Munddurchmesser ( rund ): unbekannt
Dorsale Schuppen: 25 Reihen mit je 40-45 rhombischen Stielschuppen mit seitlichen und medianen Kielen (4x6 µm); Stiele 6-7 µm; Endplatten der letzten Endreihen oft vergrößert (12-17 µm); Hinterende frei; zwischen Zehen 4 kurze Stacheln auf ovalen Schüppchen
Ventrale Schuppen: vZwf mit Stielschuppen bedeckt
Ökologie: Faulschlammbewohner
Ähnliche Arten: unverkennbar, variable Art
Besonderheiten: 3 Zähne, Eier glatt (125x74 µm)
Aspidiophorus paradoxus ist mit ca. 300µm Länge die größte Aspidiophorus-Art.
Dorsale Schuppen
Das gesamte Tier ist mit relativ großen rhombischen Stielschuppen bedeckt.
Querschnitt
Im Querschnitt ist der Aufbau der Stielschuppen gut zu erkennen: die Schuppen sitzen mit einer kleinen Basisplatte auf der Kutikula der Tiere, von der sich ein dünner, hohler Stiel erhebt. Am Ende des Stiels sitzt eine rhombische Endplatte, die mit einem Mittelkiel versehen ist. Am hinteren Ende des Tieres sind die Endplatten der letzten Schuppenreihe vergrößert.
Der Pharynx der Tiere ist terminal geschwollen, der Kopf ist schwach fünflappig mit zwei getrennten Paaren Tasthaar-Büscheln.
Ventrale Ansicht
Ventral fällt das starke Hypostomion hinter der Mundöffnung auf. Die beiden Wimpernbänder spalten sich am Kopf auf, die inneren Äste vereinigen sich bei der von mir untersuchten Population aber nicht. Die Zehenbasis trägt keine Schuppen, die Haftröhrchen
messen etwa 50% bis 70 % der Zehenlänge und laufen spitz zu.
Sehen wir uns die Schuppen etwas genauer an:
Rückenschuppen
Bei den Schuppen am Rücken erkennt man die rhombische Form der Endplatten am deutlichsten. Weniger auffällig - aber arttypisch - ist der Mittelkiel der Schuppen.
Querschnitt Schuppen
Im Querschnitt wird die komplexe Geometrie der Stielschuppen deutlich - Basisplatte, Stiel und Endplatte bilden einen sehr flexiblen und stabilen Panzer. Der zusätzliche Hohlraum unter den Außenschuppen wirkt wie eine “Knautschzone” und erhöht die Schutzwirkung zusätzlich.
Querschnitt Schuppenstiele
Die Stiele der Schuppen bestehen aus hohlen Röhrchen, die maximale Stabilität bei minimalem Materialaufwand gewährleisten - ein faszinierendes Beispiel für evolutionäre Optimierung.
Der Hinterleib der Tiere ist nicht vollständig mit Stielschuppen bedeckt. Vielmehr enden sie in der Aftergegend und es schließen sich einfache kleine, rundliche Kielschuppen an, die kein Hindernis für den Kot darstellen.
Beschuppung des Hinterleibs
An der Furca-Basis ragen einige (lt. Literatur 4) Stacheln in den Zehenausschnitt.
Der Kopf ist nahezu vollständig mit etwas kleineren Stielschuppen bedeckt:
Kopfschuppen
Kephalion und Pleuren sind recht klein und unscheinbar.
Laut Literatur besitzt A. paradoxus drei Zähne im Pharynx:
Mundbewaffnung
In den von mir untersuchten Tieren war lediglich eine gebogene Stilett-Spange zu finden, deren Spitze in das Lumen des Pharynx hineinragt und wohl zum Öffnen von Algenzellen dient, die an ihr vorbei gefördert werden. Möglicherweise beruht die Literaturangabe “drei Zähne” lediglich auf ein mikroskopisches Artefakt, da die gesamte Spange unter Umständen nicht als Ganzes in der Fokusebene liegt.
Dorsale Schuppen: 45-53 Reihen zu je 29-32 Stielschuppen (5-7 µm); kein Kiel, nicht rhombisch, ähnliche Form wie bei
P. rhomboides
; Zehenbasis dorsal mit Kielen
Ventrale Schuppen: 6 gekielte Terminalplatten; am Hinterende 10-12 Reihen von Kielen (4-4.5 µm), sonst nackt
Ökologie: zwischen Wurzeln; Brasilien
Besonderheiten: sehr kleiner Pharynx; Körper sehr breit
Dorsale Schuppen: 18-20 Reihen mit je 42-45 kleinen Stielschuppen; Stiele 1.5 µm; Endplatten 2-3.5 µm lang, proximal verstärkter Rand, distal zugespitzt, schwacher Mittelkiel; keine Stacheln
Ventrale Schuppen: vZwf. nackt, bis auf wenige längliche Stielschuppen; zwei schmale Terminalplatten(6.5-11.5 µm)
Ökologie: Moor, Schlamm
Ähnliche Arten: abgegrenzt durch nacktes vZwf
Besonderheiten: ohne Pseudozellen; nacktes vZwf.
Diesen Bauchhärling habe ich bisher nur einmal in einem Moor gefunden (Sima-Moor). Durch das nackte ventrale Zwischenfeld mit langen Terminalplatten ist die Art gut abzugrenzen:
Gut sind auch die runden Vorderränder der Endplatten der lateralen Stielschuppen zu erkennen. Das gefundene Tier ist mit ca. 130 µm etwas kleiner als die Literaturangabe und alle anderen Maße skalieren entsprechend. Der Mundring ist sehr klein und stark ausgeprägt. Ein Hypostomium existiert nicht.
Im medianen optischen Schnitt ist die hantelförmige Pharynx-Form gut zu erkennen. Die Schuppen sind distal lange ausgezogen (leider war die Form der Endplatten nicht zu erkennen). Interessant ist auch die körnige Struktur der beiden Klebedrüsen.
Dorsale Schuppen: 12-15 Reihen mit je 40-42 Stielschuppen mit rhombischen Endplatten ohne Mittelkiel (3-4 x 5-6 µm); Schuppenpanzer endet in Afterregion; Hinterende und Zehen nackt; 3 Paar kurze Stacheln an Zehenbasis (8-10 µm)
Ähnliche Arten: A. paradoxus
: ungekielte Basisplatten; Bestachelung am Hinterende; Mundbewaffung
Besonderheiten: Mundbewaffnung spitze Spange
Von den (lt. (Schwank, 1990))
zwanzig limnischen Aspidiophorus-Arten sind in den “offiziellen” Artenlisten für Deutschland lediglich vier aufgeführt. Eine weitere Art wurde vor einiger Zeit von Michael Plewka beschrieben. Die Art Aspidiophorus squamulosus wurde bisher lediglich in Polen und in Westfrankreich nachgewiesen - also kein Wunder, dass dieses Tier auch bei uns, zwischen den bisherigen Fundgebieten, beheimatet ist.
Bild 1: Aspidiophorus squamulosus; optischer Schnitt und Schuppenbild
Der innere Aufbau des Tieres wird in der Seitenansicht recht deutlich:
Bild 2: A. squamulosus, Seitenansicht
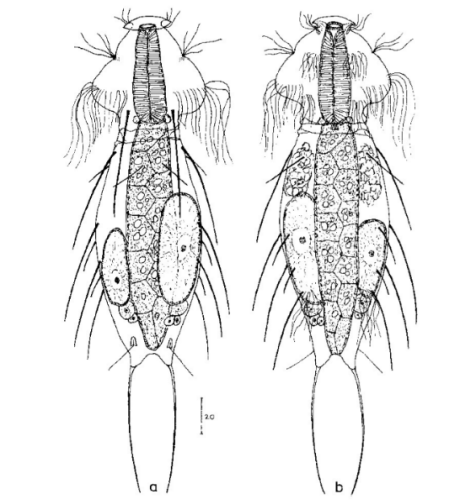
A. squamulosus ist mit ca. 230µm etwas kleiner als die ähnliche Art A. paradoxus und grenzt sich eindeutig durch die je drei Stacheln über den Zehen ab (Bild 3, ob. li. und mi. re.). Außerdem besitzen die Endplatten der Stielschuppen keinen Mittelsteg. Auch das prominente, gefurchte Hypostomium (Bild 3, ob. re.) ist typisch für diese Art. Einige von mir gefundenen Tiere waren in ihrer “Zwitterphase” (post-parthenogenetische Phase) und zeigten ein sehr großes X-Organ und Spermienbündel (Bild 3, mi. li.). Ein Tier trug zusätzlich ein sehr große Ei (mit einem ausgeprägtem Nukleolus), das es im Laufe der ca. 10-tägigen Beobachtungszeit abgelegte. Leider habe ich die Eiablage selbst nicht beobachten können (auch das Ei habe ich im Präparat nicht gefunden).
Bild 3: A. squamulosus: Details; ob. li.: Zehenstacheln; ob. re.: Hypostomium mi. li.: X-Organ und Spermien (Markierung); mi. re.: Zehenstachel und Zehenschuppen; un. li.: Querschnitt Schuppenpanzer; un. re.: Stilette (Markierung)
Wie so oft gab es einige Abweichungen von der Artbeschreibung in der Literatur. Bei bis zu 50 Generationen pro Jahr und jahrelanger Isolation in einen kleinen Gartenteich finde ich kleinere Abweichungen nicht verwunderlich. Möglicherweise sind aber auch bei den seltenen, früheren Beobachtungen einige Details nicht aufgefallen. So sollte die Zehenbasis unbeschuppt sein (Bild 3, mi. re.). Die von mir beobachteten Tiere zeigen aber viele sehr kleine, rundliche und enganliegende Schuppen. Lediglich die auffallenden Stielschuppen kommen am Hinterende des Tieres nicht mehr vor.
Bei einigen Aspidiophorus-Arten sind bewegliche “Zähne” in der Mundröhre beschrieben. Obwohl A. squamulosus zahnlos sein sollte, konnten zwei aktiv bewegliche Stilette beobachtet werden, deren Spitze in der Mundröhre endeten und an Tardigraden erinnerten (Bild 3, un. re.).
Die eigenartigen Stiel-Schuppen der Gattung Aspidiophorus sind im Schuppenverband nicht in ihrer dreidimensionalen Schönheit zu erfassen. Deshalb habe ich ein bedauernswertes Exemplar von A. squamulosus einer Schuppenanalyse unterzogen:
Bild 4: A. squamulosus; gefärbte und - leider - platt gedrückte Einzelschuppen
Da bei einer Schuppenanalyse gerade die sehr kleinen Schuppe ziemlich platt gedrückt werden, habe ich mich mal an einer zeichnerischen Rekonstruktion der dreidimensionalen Struktur versucht:
Bild 5: A. squamulosus; unten: Schuppenbild im Verbund; links oben: vereinzelte und gefärbte Schuppen; rechts oben: zeichnerische Rekonstruktion einer einzelnen Stiel-Schuppe
Ein weitere Punkt, der bisher offen blieb, ist der Speiseplan von A. squamulosus. Die Hauptnahrung besteht aus relativ großen einzelligen Algen bzw. Augenflagellaten. Um diese große Beute zu verschlingen, ist der Mund der Tiere mit beweglichen Lamellen ausgestattet, die zur Nahrungsaufnahme nach außen geklappt werden können (vgl. Captochaetus). Dadurch ist der Durchmesser der Mundöffnung - anders als bei den meisten Gastrotrichen - stark erweiterbar. Bei der Schluckbewegung werden die Stilette in die Mundröhre gedrückt und die Beutezellen an ihnen vorbei geführt. Dabei wird die harte Zellwand wohl perforiert, so dass die Verdauungssekrete im Darm in die Zellen eindringen können.
Bild 6: A. squamulosus; mit Augenflagellaten gefüllter Darm
Untersucht man den Schlamm von stehenden Gewässern mikroskopisch, findet man immer wieder Gastrotrichengelege in leeren Schalen von Wasserflöhen mit manchmal dutzenden Eieren.
Bild 7: A. squamulosus; Fokusstack eines Gastrotrichengelege mit Eieren von mindestens drei unterschiedlichen Arten
Oft bestehen diese Gelege aus Eiern unterschiedliche Arten. Anscheinend legen eine Reihe von Arten ihre Eier bevorzugt in Wasserfloh-Schalen, in denen bereits andere Gastrotrichen ihre Eier abgelegt haben. In einem solchen Gelege fallen oft relativ große (95µm x 55 µm), bestachelte Eier auf, die noch nicht mit der Embryonalentwicklung begonnen haben und wohl Dauereier darstellen.
Bild 5: A. squamulosus; Dauerei
Ein Ei des gezeigten Geleges begann sich zu entwickeln und stellte sich als ein Ei der Art A. sqamulosus heraus. Deshalb kann ich hier die zur Art gehörigen Eier zeigen.
Dorsale Schuppen: 15-20 Reihen mit je 38-40 sehr kleinen Stielschuppen; Endplatten sehr zart und ungekielt (2.5-3 µm); stark gebogene Stiele -> Schuppen stehen borstenartig ab
Ventrale Schuppen: vZwf. 9-10 Reihen Kiele; vorne meist nackt; keine Terminalplatten
Ökologie: Bach
Ähnliche Arten: gut abgrenzbar durch abstehende Schuppen
Dorsale Schuppen: 15-17 Reihen mit je 33-40 Stielschuppen; Endplatten ungekielt 2-5µm, distal und proximal abgerundet; Zehenbasis nackt; 2 Paar dicke Stacheln (8-10µm und 4-5µm) auf ovalen Schuppen mit zwei Kielen
A. tetrachaetus ist ein sehr seltener Gastrotrich, der bisher nur für Polen bestätigt war.
A. tetrachaetus: dorsal ((c) Ole Riemann)
Der Rücken der Tiere ist mit relativ großen, ungekielten und abgerundeten Stielschuppen bedeckt, die sich am Hinterende nicht bis zur Zehenbasis fortsetzen. Am Hinterende des Schuppenkleides fällt ein Paar großer Stacheln auf.
A. tetrachaetus: Querschnitt ( Ole Riemann)
Im Querschnitt erkennt man gut den Aufbau der Stielschuppen.
A. tetrachaetus: ventral (copyright: Ole Riemann)
Das ventrale Zwischenfeld der Tiere ist lediglich in der Darmregion von 8-9 Längsreihen kleiner Kiele bedeckt. Die Pharynx-Region ist nackt. Zwei terminale Kiele bilden den hinteren Abschuss der Bauchbeschuppung.
A. tetrachaetus: zwei Stachelpaare am Hinterende
Erst ein genauer Blick auf die dorsale Zehenbasis der Tiere lässt zwei zusätzliche, kleinere Stacheln erkennen.
A. tetrachaetus: einzelner Zahn in der Mundhöhle
Entgegen der Beschreibung in der Literatur, ist in bei den Tieren im Sima-Moor ein einzelner Zahn zu erkennen, der unsymmetrisch aus der Mundhöhle ragt.
Schuppe trägt einen Stachel, der sich an der Basis in zwei gleich lange Äste spaltet
Einschnürung an der Furca-Basis
Arten
2 Arten:
Bifidochaetus articus Dorsal: 36-45 Reihen mit je 30-42 dreickig gerundete, nicht überlappende Schuppen; Schuppen tragen einen gespaltenen Stachel (5-11 µm) Ventral: vZwf. nackt bis auf Hinterende; hier 3-5 Reihen zu je 5 ovalen Schuppen; zwei Terminalplatten
Bifidochaetus veronicae Dorsal: 16-19 Reihen mit je 27-30 subovale überlappende Schuppen (3x3 µm) mit gespaltenem Stachel (9µm); ein Paar parafurcale Stacheln (14-16 µm) Ventral: vZwf. nackt bis auf hinteres Drittel, das kleine, ovale Kiele (2x2 µm) trägt; zwei gekielte Terminalplatten (6x4 µm); 7-8 terminale Stacheln (10-14 µm)
Dorsale Schuppen: 16-19 Reihen mit je 27-30 subovale überlappende Schuppen (3x3 µm) mit gespaltenem Stachel (9µm); ein Paar parafurcale Stacheln (14-16 µm)
Ventrale Schuppen: vZwf. nackt bis auf hinteres Drittel, das kleine, ovale Kiele (2x2 µm) trägt; zwei gekielte Terminalplatten (6x4 µm); 7-8 terminale Stacheln (10-14 µm)
Cephalionotus kisielewskii Besonderheiten: dreiteilige Haftröhrchen Dorsal: 7-9 Reihen zu je 17-18 unbestachelte Schuppen; vordere 2/3 rund bis hexagonal, letzte 5 Reihen je 3-5 größere Schuppen Ventral: großes Hypostomium, vZwf. mit 3-5 Reihen mit je 15-17 stachellosen, rechteckigen Schuppen; 2 große gekielte Terminalplatten, gefolgt von einem Paar schmaler, langer Kiele
Haftröhrchen (dreiteilig (17, 10, 7 µm)): 80% der Furka
Pharyx ( hantelförmig ):
Munddurchmesser ( rund ): unbekannt
Dorsale Schuppen: 7-9 Reihen zu je 17-18 unbestachelte Schuppen; vordere 2/3 rund bis hexagonal, letzte 5 Reihen je 3-5 größere Schuppen
Ventrale Schuppen: großes Hypostomium, vZwf. mit 3-5 Reihen mit je 15-17 stachellosen, rechteckigen Schuppen; 2 große gekielte Terminalplatten, gefolgt von einem Paar schmaler, langer Kiele
Ökologie: Flußbett, sandig und schlammig, Brasilien
Chaetonotus bogdanovii Dorsal: 5 Reihen mit je 20 einfachen, dicken und kurzen Stacheln; lateral 12 Großstacheln: 2 Paar am Hals, 1 Paar in der Mitte, 3 Paar am Hinterende Ventral:
Chaetonotus (C. ) hoanicus Dorsal: Kopf: 18-19 Reihen; Rumpf 10-11 Reihen mit insgesammt je 33-40 dünner, sich stark überlappender siebeneckiger bis rechteckiger Schuppen (19µm x 9µm), die einen Stachel (2µm - 40µm) mit Nebenspitze tragen; kurz bestacheltes Feld am Hinterende Ventral: ventrales Zwischenfeld mit zwei gebogenen, gekielten Terminalplatten;8-9 Reihen Kielschuppen (zum Teil bestachelt); in der Pharynx-Region ovale, glatte Schuppen
Chaetonotus (C. ) multispinosus Dorsal: 17-19 Reihen mit je 38-40 kreisrunden Schuppen mit distalem Ausschnitt und kurzen und einfachen Stacheln Ventral: 2-3 Paar länglicher, gekielter Terminalplatten; 5-7 Reihen kleine, rund Kielschuppen
Dorsale Schuppen: Kopf: 18-19 Reihen; Rumpf 10-11 Reihen mit insgesammt je 33-40 dünner, sich stark überlappender siebeneckiger bis rechteckiger Schuppen (19µm x 9µm), die einen Stachel (2µm - 40µm) mit Nebenspitze tragen; kurz bestacheltes Feld am Hinterende
Ventrale Schuppen: ventrales Zwischenfeld mit zwei gebogenen, gekielten Terminalplatten;8-9 Reihen Kielschuppen (zum Teil bestachelt); in der Pharynx-Region ovale, glatte Schuppen
Besonderheiten: Ausgeprägtes System von Pharynx-Adern
Chaetonotus hoanicus ist ein sehr seltener Gastrotrich, der bisher nur von Schwank beobachtet wurde. Deshalb möchte ich ihn etwas näher beschreiben. Das Hauptmerkmal von Ch. hoanicus ist der abrupte Übergang von einem unbestachelten Kopfbereich zu einem relativ lang bestachelten Rücken:
Ch. hoanicus: Änderung der dorsalen Bestachelung
Die Beschuppung des Hals- /Kopfbereichs besteht aus nahezu runden, nur mit einem extrem kurzen - oft nur erahnbaren - Stachel, versehenen stark überlappenden Schuppen:
Ch. hoanicus: Änderung der dorsalen Beschuppung am Hals
Mit einem abrupten Übergang zu lang bestachelten Schuppen wird der Bereich der Rumpfbeschuppung deutlich abgetrennt. Die Rumpfschuppen sind relativ groß, grob fünf eckig und haben einen großen Ausschnitt am Hinterende. An dessen Spitze entspringen ca. 15-20 µm lange Stacheln, die distal mit einer Nebenspitze versehen sind.
Ch. hoanicus: Änderung der dorsalen Rumpf-Bestachelung
In der Afterregion schließt sich ein kurzbestacheltes Feld an. In den Zehenausschnitt ragen einige auffällige, längere Stacheln.
Ch. hoanicus: Änderung der dorsalen Bestachelung der Afterregion
Die genaue Schuppenform wird erst deutlich, wenn es gelingt, Schuppe abzulösen:
Ch. hoanicus: Dorsale Einzelschuppen aus dem Rückenfeld
Auffällig ist die besonders ausgeprägte stimmgalbelförmige Strukturierung der Pharynxoberfläche, die die Mundhöhle mit den Drüsen ind der Pharynx-Wand (siehe Pharynx) verbindet:
Ch. hoanicus: Struktur auf Pharynx-Oberfläche
Filmt man ein lebendes Exemplar von Ch. hoanicus mit dem Fokus auf diese “Adern”, wird eine deutliche Strömung innerhalb dieser Strukturen sichtbar, die auf eine drüsenartige Funktion hindeuten:
Der kleine und häufige Gastrotrich Chaetonotus (Ch.) multispinosus ist durch die kurzbestachelten, rundlichen und am Hinterende ausgeschnittenen Schuppen gekennzeichnet:
Ch multispinosus: ventral
Im Querschnitt erkennt man die namesgebenden kurzen Stacheln. Sehr typisch ist der kurze, keulenförmige Pharynx:
Ch multispinosus: Querschnitt
Das ventrale Zwischenfeld ist mit wenigen reihen rundlichen, gekielten Schuppen bedeckt. Den hinteren Abschluss bilden zwei größere, kurz bestachelte Platten , die von zwei Paaren kleiner, ebenfalls kurz bestachelter Kielschuppen eingefasst sind:
Für die Untergattung Hystricochaetonotus sind die relativ großen, sich nicht überlappenden Schuppen mit einem langen Stachel. Diesem stacheligen Aussehen verdankt diese Untergattung ihren Namen (hystix = Stachelschwein)
Die Schuppen stehen in typischer Anordnung. Ihre Form ist dreilappig mit einem Mittelkiel, an dessen Ende sich ein langer - meist mit Nebenspitze versehenen - Stachel entspringt:
typische dreilappige Schuppen bei Hystricochaetonotus - hier
C. (H. ) persetosus
Diagnose
Körperlänge 60 - 190 µm
dreilappige Schuppen mit axialem Kiel
Stachel mit Nebenspitze (kann aber manchmal auch fehlen oder doppelt sein)
alle Schuppen bestachelt, Stacheln verlängern sich von vorne nach hinten
Chaetonotus (H. ) italicus Dorsal: 13 Reihen mit je 13-14 dreilappigen Schuppen (6,5µm x 5µm) mit einfachem Stachel (3µm - 10µm); für Hystricochaetonotus typische Schuppengruppe ohne Stacheln am Hinterende Ventral: ventrales Zwischenfeld mit zwei Reihen mit je 20 rechteckige Schuppen (1,2µm - 4µm); zwei gekielte Terminalplatten (Länge 6,4µm - 6,8µm)
Chaetonotus (H. ) murrayi Dorsal: 10-11 Reihen mit je 19-21 dreilappigen Schuppen (6µm - 11µm) mit langen, zweispitzigen Stacheln (20-23 µm), die fast senkrecht vom Körper starr abstehen Ventral: 2 lange, dünne Terminalplatten mit kurzen Stacheln; 7-8 Reihen mit kleinen Kielschuppen (1µm - 3µm)
Chaetonotus (H. ) novenarius Dorsal: 7 Reihen am Kopf, 12 am Rumpf mit je 14-15 dreilappigen, gekielten Schuppen (3-6 µm), die nicht überlappen und kurz und zart bestachelt sind (10µm); am Kopfende zwei Reihen mit längeren Stacheln. Auf einem relativ engen Feld am Vorderrumpf sehr lange Stacheln mit zwei Nebenspitzen (26-50µm) Ventral: 2 länglich-ovale, gekielte Terminalplatten (6,6µm x 4µm) mit zwei einfachen Stacheln; 4-6 Reihen mit je 40 ovalen Kielschuppen. Eigene Beobachtung: in Pharynxgegend rechteckige Querspangen
Dorsale Schuppen: 13 Reihen mit je 13-14 dreilappigen Schuppen (6,5µm x 5µm) mit einfachem Stachel (3µm - 10µm); für Hystricochaetonotus typische Schuppengruppe ohne Stacheln am Hinterende
Ventrale Schuppen: ventrales Zwischenfeld mit zwei Reihen mit je 20 rechteckige Schuppen (1,2µm - 4µm); zwei gekielte Terminalplatten (Länge 6,4µm - 6,8µm)
Ökologie: Moor
Ähnliche Arten: alle Hystricochaetonotus; genaue Diagnose erforderlich
Besonderheiten: Stacheln ohne Nebenspitze; einzigartige Beschuppung des ventralen Zwischenfeldes
Der bisher nur aus Italien bekannte, kleine Gastrotrich Chaetonotus (H.) italicus ist im Sima-Moor relativ häufig anzutreffen.
Ch. italicus: dorsal
Viele der gefundenen Exemplare waren in ihrer postparthenogenetischen (Zwitter-) Phase und trugen sowohl ein deutliches, zweiteiliges X-Organ als auch stäbchenförmige Spermatoiden. Typisch für die Art ist der ausgeprägt hantelförmige Pharynx:
Ch. italicus: Querschnitt
Das ventrale Zwischenfeld ist mit zwei Reihen rechteckiger Schuppenplatten besetzt, die im Halsbereich undeutlicher werden. Den Abschluss bilden zwei gekielte Terminalplatten:
Ch. italicus: ventral
Obwohl die Schuppenstacheln keine Nebenspitze tragen, wird die Einordnung in der Untergattung Hystricochaetonotus wegen der typischen Schuppenform deutlich:
Dorsale Schuppen: 10-11 Reihen mit je 19-21 dreilappigen Schuppen (6µm - 11µm) mit langen, zweispitzigen Stacheln (20-23 µm), die fast senkrecht vom Körper starr abstehen
Ventrale Schuppen: 2 lange, dünne Terminalplatten mit kurzen Stacheln; 7-8 Reihen mit kleinen Kielschuppen (1µm - 3µm)
Ökologie: Moor, Ufermoos
Besonderheiten: Zehen nackt; starr abstehende Stacheln sind sehr typisch
Der sehr seltene Gastrotrich Chaetonotus (H.) murrayi ist bisher nur von zwei Fundstellen (in Polen und bei Berlin) bekannt. Die starken und recht langen zweispitzigen Stacheln stehen spießförmig ab und geben dem Tier ein sehr wehrhaftes Aussehen.
Ch. murrayi: Querschnitt
Besonders auffällig wird das beim Fokus auf die Rückenbestachelung:
Ch. murrayi: dorsal
Die ca. 20µm langen Stacheln tragen nahe der Hauptspitze eine laterale Nebenspitze:
Ch. murrayi: Fokus auf die Stachelspitzen
Der Fokus auf die Schuppen zeigt die typische dreilappige Schuppenform dieser Untergattung. Abweichend zu den Funden in der Literatur ist auch der gesamte Kopf der Tiere mit langen Stacheln besetzt:
Ch. murrayi: dorsale Beschuppung des Kopf / Halsbereichs
Die Bestachelung ist auch Hinterende deutlich ausgeprägt. Anders als viel Mitglieder der Untergattung Hystrocochaetonotus fehlen hier die unbestachelten dorsalen Kielplatten:
Ch. murrayi: dorsale Beschuppung des Hinterendes
Das ventrale Zwischenfeld trägt 6 - 7 Reihen kleiner Kielschuppen und wird durch zwei bestachelte Terminalplatten abgeschlossen.
Dorsale Schuppen: 7 Reihen am Kopf, 12 am Rumpf mit je 14-15 dreilappigen, gekielten Schuppen (3-6 µm), die nicht überlappen und kurz und zart bestachelt sind (10µm); am Kopfende zwei Reihen mit längeren Stacheln. Auf einem relativ engen Feld am Vorderrumpf sehr lange Stacheln mit zwei Nebenspitzen (26-50µm)
Ventrale Schuppen: 2 länglich-ovale, gekielte Terminalplatten (6,6µm x 4µm) mit zwei einfachen Stacheln; 4-6 Reihen mit je 40 ovalen Kielschuppen. Eigene Beobachtung: in Pharynxgegend rechteckige Querspangen
Ökologie: Moor, pflanzenreiche Kleingewässer
Ähnliche Arten: C. (H. ) anomalus
ist möglicherweise synonym; unterscheidet sich nur durch das Fehlen der beiden Schuppenreihen mit längeren Stacheln am Kopf (die bei meinen Beobachtungen vorhanden war) und durch Schuppenspangen unter dem Pharynx (die bei meinen Beobachtungen auch bei Ch. novenarius vorhanden sind)
Besonderheiten: durch die langen Stacheln, die aus einem engen Feld entspringen, gut abgrenzbar
Chaetonotus (H.) novenarius fällt sofort durch seine neun sehr langen Großstacheln auf.
Ch. novenarius: dorsal
Die neun Großstacheln tragen jeweils zwei Nebenspitzen und überragen das Zehenende des Tieres. Ch. novenarius ist der Chaetonotus mit den relativ zur Körperlänge längsten Stacheln (ca. 2/3 der Körperlänge):
Ch. novenarius: dorsale Großstacheln
Obwohl die Beschuppung und Bestachelung das Tier eindeutig als Ch. novenarius ausweisen, gibt die ventrale Beschuppung der gefundenen Tier Rätsel auf:
Ch. novenarius: ventral
Auffallend sind die rechteckigen Schuppenplatten im Pharynx-Bereich, die Schwank nur für die sehr ähnliche Art Ch. anomalus aufweist. Möglicherweise handelt es sich bei dieser Art - wie bereits (Schwank, 1990)nicht ausschließt - um ein Synonym der Art Ch. novenarius.
dicke, gerade Stacheln am Hinterende der Schuppen (selten gebogen)
alle Stacheln +/- gleich lang, Länge nimmt nach hinten leicht zu
manchmal Gruppen mit verlängerten Stacheln
Arten
21 Arten:
Chaetonotus (P. ) acanthocephalus Dorsal: 5 Reihen mit je 15-18 querovalen Schuppen (5 x 10 µm), die sich nicht berühren; am Kopf dorsal und lateral 5-7 charakteristische Stachelschuppen; kleine Stacheln 6-7 µm; Gruppen von Großstacheln 8-22 µm Ventral: Ventrales Zwischenfeld am Kopf 3 sich berührende Platten; sonst elliptische Platten; in Darmregion manchmal 3 Rehen von kleineren elliptischen Platten
Chaetonotus (P. ) acanthodes Dorsal: 10 Reihen mit je 20-25 stark überlappende, abgerundet dreickige Schuppen (5 µm - 8.5 µm) mit stark aufgewölbten Rand (Rand erscheint doppelt) mit kurzen Stacheln (7 µm); 7 - 12 einfache Gürtelstacheln (6.5 µm - 16.5 µm) in alternierenden Reihen; Schuppen hinter Stachelgürtel meist unbestachelt Ventral: 4 längliche Terminalplatten mit Kiel; 8-9 Reihen kleiner Kielschuppen
Chaetonotus (P. ) cordiformis Dorsal: 7 Reihen mit fe 20 dünnen, querovalen bis verkehrt herzförmigen Schuppen, die sich nur wenig berühren; Stacheln (10-39 µm) entspringen etwas hinter Schuppenmitte; Nebenspitze relativ weit vom distalen Ende entfernt; drei Paar lange laterale Stacheln (34-46 µm) überragen die Zehen um das doppelte ihrer Länge; zwischen den Zehen 3 dorsale Stacheln, die die Zehen etwas überragen Ventral: Ventrales Zwischenfeld mit +11 Reihen kurz bestachelter winzigen Schuppen
Dorsale Schuppen: 5 Reihen mit je 15-18 querovalen Schuppen (5 x 10 µm), die sich nicht berühren; am Kopf dorsal und lateral 5-7 charakteristische Stachelschuppen; kleine Stacheln 6-7 µm; Gruppen von Großstacheln 8-22 µm
Ventrale Schuppen: Ventrales Zwischenfeld am Kopf 3 sich berührende Platten; sonst elliptische Platten; in Darmregion manchmal 3 Rehen von kleineren elliptischen Platten
Ökologie: Moorgewässer, selten, sehr variabel
Besonderheiten: sehr variabel bezüglich Stachelgröße - möglicherweise eine Reihe von Unterarten
Ein sehr seltener und seltsamer Bauchhärling ist der nahezu drehrunde Chaetonotus (P.) acanthocephalus mit seiner unregelmäßigen Bestachelung und den großen, runden Schuppen:
Ch. acanthocephalus: Fokus auf das dorsale Schuppenkleid
Ventral trägt das Tier am Kopf große kutikulare Platten die sich zum Hinterende hin in runde, kurzbestachelte Schuppen wandeln:
Ch. acanthocephalus: ventral
Mazeriert man das Tier mit Eisessig, wird die für die Untergattung Primochaetus typischen Schuppenform besonders deutlich:
Ähnliche Arten: durch Stachelgürtel und doppeltem Schuppenrand gut abgrenzbar
Besonderheiten: Stachelgürtel und doppeltem Schuppenrand
Ein weiterer Vertreter der Untergattung Primochaetus ist der kleine Bauchhärling Ch. (P.) acanthodes, der durch seine deutlichen, stachellosen Schuppen mit “doppelter” Vorderkontur und einem Gürtel langer Stacheln am Hinterleib auffällt:
Ch. acanthodes: dorsale Schuppen
Erst in der Seitenansicht erkennt man, dass der Stachelgürtel durchgängig die Rückenseite des Tieres umfasst:
Ch. acanthodes: Seitenansicht
Das ventrale Zwischenfeld ist mit ca. 8 Reihen kleiner, rundlicher Kielschuppen bedeckt. Den hinteren Abschluss bilden vier rechteckige, schmale Kielplatten:
Ch. acanthodes: ventrale Beschuppung
Im Querschnitt wird klar, dass die Schuppen stark gebogen sind. Dadurch entsteht die “Doppelkontur” im o0ischen Schnitt. Am Pharynxeingang sitzen bei den von mir gefundenen Tieren ein bis zwei Zähne:
Dorsale Schuppen: 7 Reihen mit fe 20 dünnen, querovalen bis verkehrt herzförmigen Schuppen, die sich nur wenig berühren; Stacheln (10-39 µm) entspringen etwas hinter Schuppenmitte; Nebenspitze relativ weit vom distalen Ende entfernt; drei Paar lange laterale Stacheln (34-46 µm) überragen die Zehen um das doppelte ihrer Länge; zwischen den Zehen 3 dorsale Stacheln, die die Zehen etwas überragen
Ventrale Schuppen: Ventrales Zwischenfeld mit +11 Reihen kurz bestachelter winzigen Schuppen
Besonderheiten: Brutpflege: dekorieren der Eier mit Detritus; var. bernensis hat keine Nebenspitzen
Ch. cordiformis zeigt im Querschnitt den typischen Aufbau limnischer Gastrotrichen. Auffallend sind die relativ wenigen, langen und stark gebogenen Schuppenstacheln:
Ch. cordiformis: Querschnitt
In der Seitenansicht zeigt sich die enorme Länge der mit einer Nebenspitze versehenen Stacheln:
Ch. cordiformis: Seitenansicht
Diese langen Stacheln erschweren es sehr, gute Fotos von den Tieren zu machen, da das Festlegen der Tiere mit dem Deckglas die natürliche Anordnung der Stacheln stark stört. Deshalb sollte versucht werden, das Erscheinungsbild der Tiere freischwimmend zu erfassen:
Ch. cordiformis: freischwimmendes Exemplar mit Fokus auf die Schuppenstacheln
Hochauflösende Aufnahmen werden erst bei etwas gepressten Tieren möglich. Erst hier wird die rundliche Schuppenform mit den mit einer Nebenspitze versehenen Schuppenstacheln sichtbar. Die Nebenspitze sitzt relativ weit von dem Stachelende der Tiere entfernt.
Ch. cordiformis: Stachelschuppen mit Nebenspitze
Viele der gefundenen Exemplare wiesen eigenartige kugelförmige Körper im Inneren auf, die ich für (Dauerformen? von) Parasiten halte:
Ch. cordiformis: kugelförmige Einschlüsse
Die Schuppen der Tiere sind erst nach dem Ablösen gut beurteilbar. Hier erkennt man die namesgebende “Herzform” der rundlichen Schuppen mit Einbuchtung:
Ch. cordiformis: herzförmige Schuppen
Ch. cordiformis ist einer der wenigen Gastrotrichen, der nach der Eiablage eine Art “Brutpflege” betreibt. Nach der Eiablage transportiert die Mutter Detritus aus der Umgebung des Ablageortes in Ihrem Pharynx zum EI und klebt dieses Material an die Eischale. Im Ergebnis werden die Eier vollständig von Detritus bedeckt und durch ihn vor Feinden geschützt:
Ch. cordiformis: Brutpflege - Film durch anklicken starten
Dorsale Schuppen: 5 Reihen mit je 20 einfachen, dicken und kurzen Stacheln; lateral 12 Großstacheln: 2 Paar am Hals, 1 Paar in der Mitte, 3 Paar am Hinterende
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Schimkewitsch,W. 1886. Uber eine neue Spezies Ichthydium. Nachr. Gesell. Freunde Naturw. Moskau 50: 148-150.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus crinitus
Taxonomie
Ordnung: CHAETONOTIDA Remane, 1925
UnterOrdnung: PAUCITUBULATINA d'Hondt, 1971
Familie: CHAETONOTIDAE Gosse, 1864
Unterfamilie: CHAETONOTINAE Kisielewski, 1991
Gattung: Chaetonotus Ehrenberg, 1830
Art: crinitus
Länge ( flaschenförmig ):
Breite:
Kopfbreite ( fünflappig ):
µm
Furkalänge:
Haftröhrchen:
Pharyx ( zylindrisch ):
Munddurchmesser ( rund ): unbekannt
Besonderheiten: unzureichend beschrieben und so nicht wieder zu erkennen!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Sudzuki,M. 1971. The Gastrotricha of Japan which live in the capillary water of the interstitial system: II. Bull. Biogeogr. Soc. Japan 27: 37-41.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus dentatus
Taxonomie
Ordnung: CHAETONOTIDA Remane, 1925
UnterOrdnung: PAUCITUBULATINA d'Hondt, 1971
Familie: CHAETONOTIDAE Gosse, 1864
Unterfamilie: CHAETONOTINAE Kisielewski, 1991
Gattung: Chaetonotus Ehrenberg, 1830
Art: dentatus Tretyakova, 1992
Länge ( flaschenförmig ):
Breite:
Kopfbreite ( fünflappig ):
µm
Furkalänge:
Haftröhrchen:
Pharyx ( zylindrisch ):
Munddurchmesser ( rund ): unbekannt
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Chaetonotus elegantulus
Taxonomie
Ordnung: CHAETONOTIDA Remane, 1925
UnterOrdnung: PAUCITUBULATINA d'Hondt, 1971
Familie: CHAETONOTIDAE Gosse, 1864
Unterfamilie: CHAETONOTINAE Kisielewski, 1991
Gattung: Chaetonotus Ehrenberg, 1830
Art: elegantulus Remane, 1936
Länge ( flaschenförmig ):
Breite:
Kopfbreite ( fünflappig ):
µm
Furkalänge:
Haftröhrchen:
Pharyx ( zylindrisch ):
Munddurchmesser ( rund ): unbekannt
Besonderheiten: sp. inquirenda; jegliche Abbildung und Beschreibung fehlt!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Remane,A. 1936. Gastrotricha, In: H.G. Bronns, ed. Klassen Ordnungen des Tierreichs, Band 4, Abteilung II, Buch I, Teil 2, Lieferungen 1-2. Akademie Verlagsgesellschaft, Berlin.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus gracilis
Taxonomie
Ordnung: CHAETONOTIDA Remane, 1925
UnterOrdnung: PAUCITUBULATINA d'Hondt, 1971
Familie: CHAETONOTIDAE Gosse, 1864
Unterfamilie: CHAETONOTINAE Kisielewski, 1991
Gattung: Chaetonotus Ehrenberg, 1830
Art: gracilis Gosse, 1864
Länge ( flaschenförmig ):
Breite:
Kopfbreite ( fünflappig ):
µm
Furkalänge:
Haftröhrchen:
Pharyx ( zylindrisch ):
Munddurchmesser ( rund ): unbekannt
Besonderheiten: taxon inquirendum; keine Angaben über Schuppen und Stacheln, daher nicht wieder erkennbar
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Gosse,P.H. 1864. The natural history of the hairy-backed animalcules (Chaetonotidae). The Intellect. Observer 5: 387-406.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus hermaphroditus
Taxonomie
Ordnung: CHAETONOTIDA Remane, 1925
UnterOrdnung: PAUCITUBULATINA d'Hondt, 1971
Familie: CHAETONOTIDAE Gosse, 1864
Unterfamilie: CHAETONOTINAE Kisielewski, 1991
Gattung: Chaetonotus Ehrenberg, 1830
Art: hermaphroditus Remane, 1936
Länge ( flaschenförmig ):
Breite:
Kopfbreite ( fünflappig ):
µm
Furkalänge:
Haftröhrchen:
Pharyx ( zylindrisch ):
Munddurchmesser ( rund ): unbekannt
Besonderheiten: taxon inquirenda; jegliche Abbildung und Beschreibung fehlt!
Leider sind mir zu diese Art keine Daten zugänglich!
Wenn Sie Zugang zu folgender Literatur haben:
Remane,A. 1936. Gastrotricha, In: H.G. Bronns, ed. Klassen Ordnungen des Tierreichs, Band 4, Abteilung II, Buch I, Teil 2, Lieferungen 1-2. Akademie Verlagsgesellschaft, Berlin.
würde ich mich sehr freuen, wenn sie mir eine Kopie per Mail senden würden!
Chaetonotus lancearis
Taxonomie
Ordnung: CHAETONOTIDA Remane, 1925
UnterOrdnung: PAUCITUBULATINA d'Hondt, 1971
Familie: CHAETONOTIDAE Gosse, 1864
Unterfamilie: CHAETONOTINAE Kisielewski, 1991
Gattung: Chaetonotus Ehrenberg, 1830
Art: lancearis Tretyakova, 1992
Länge ( flaschenförmig ):
Breite:
Kopfbreite ( fünflappig ):
µm
Furkalänge:
Haftröhrchen:
Pharyx ( zylindrisch ):
Munddurchmesser ( rund ): unbekannt
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Leider sind mir zu diese Art keine Daten zugänglich!
Diesmal habe ich nicht einmal die Literaturstelle gefunden! Lediglich die Autorenangabe “Tretyakova, 1992” ist zugänglich.
Wenn Sie Zugang näher Informationen zu dieser Art haben, würde ich mich sehr freuen, wenn sie sie mir per Mail senden würden!
Chitonodytes
Diagnose
Körper flaschenförmig
Kopf durch Halseinschnürung abgesetzt, gerundet
Hinterende abgerundet oder abgestutzt
keine Schuppen
Arten
3 Arten:
Chitonodytes collini Dorsal: dorsolateral 3 Bündel mit leicht gebogenen, abspreizbaren Stacheln mit je zwei Nebenspitzen; (Hals: 3 x 130-140µm, 2 x 35-38µm; Mitte: 3 x 80-90µm; Hinten 2 x 60-70µm) Ventral: Kopf: 6 ventrolaterale Wimpernringe; Ventral 6 Wimpernbüschel; sonst nackt
Chitonodytes longisetosus Dorsal: Dorsolateral 6 Paar lange Stacheln, die das Hionterende um ca. 30µm überragen ( Hals: 3 x 80-82µm; Mitte: 2 x 60µm; Hinten: 1 x 35µm) Ventral: zwei Wimpernbänder; nackt
Chitonodytes longispinosus Dorsal: Dorsolatereal 9 Paar s-förmig geschweifte Stacheln mit 2 Nebenspritzen in 3 Bündeln (Hals: 3 x - erste zwei 180-182µm, dritte 40µm; Mitte: 4 x 130-132µm; Hinten: 2 x 60-72µm) Ventral: zwei Wimpernbänder
Dorsale Schuppen: dorsolateral 3 Bündel mit leicht gebogenen, abspreizbaren Stacheln mit je zwei Nebenspitzen; (Hals: 3 x 130-140µm, 2 x 35-38µm; Mitte: 3 x 80-90µm; Hinten 2 x 60-70µm)
Dorsale Schuppen: Dorsolateral 6 Paar lange Stacheln, die das Hionterende um ca. 30µm überragen ( Hals: 3 x 80-82µm; Mitte: 2 x 60µm; Hinten: 1 x 35µm)
Dorsale Schuppen: Dorsolatereal 9 Paar s-förmig geschweifte Stacheln mit 2 Nebenspritzen in 3 Bündeln (Hals: 3 x - erste zwei 180-182µm, dritte 40µm; Mitte: 4 x 130-132µm; Hinten: 2 x 60-72µm)
zahlreich laterale Stacheln, immer mit 1 oder 2 Nebenspitzen, die mit breiter Lamelle mit dem Hauptast verbunden sind
Stacheln der caudalen Lappen immer zweispitzig
Arten
6 Arten:
Dasydytes (D. ) asymmetricus Dorsal: Hinten: 2 stumpfe Lappen mit kurzem Stachel (17-20 µm) mit Doppel- und Nebenspitze; auf einer Seite 12, auf anderer Seite nur 3 z.T. riesige Stacheln mit Doppel- und Nebenspitze; 3 dünne Stacheln am Hinterende (50µm) Ventral: 4-5 Paar Zilienbüschel, sonst nackt
Dasydytes (D. ) goniathrix Dorsal: Auf den Hinterlappen ein Paar langer, auf Schuppenrudimenten sitzende Stacheln (63-68 µm; 1/4 Körperlänge); 2x7 Stachelbündel mit proximaler Nebenspitze; Kopf 2x mit 1 bzw 3 Stacheln; Rumpf 5x mit 4-7 Stacheln; Stachelanzahl und -länge (60.76µm) variiert Ventral: 5 Paar Wimpernbüschel
Dasydytes (D. ) monile Dorsal: Kopf zwei Ringe mit je 6 Stacheln mit einer Nebenspitze (10-15 µm); Körper 4 Paare Stachelgruppen mit je 5 Stacheln mit zwei Nebenspitzen (69-115 µm); Hinten zwei Paare Stacheln mit zwei Nebenspitzen (18µm); zwei Paar einfache Stacheln Ventral:
Dasydytes (D. ) nhumirimensis Dorsal: neun Paare Stachel-Gruppen; Stachelzahl von vorne: 1 - 1 - 2 - 5 - 5 - 3 - 1 - 1 (58-75 µm); erste beiden Gruppen einfach, sonst zwei Nebenspitzen; auf Schuppenrudimenten; Schlägerartige unpaarige Struktur am Hinterende (59x17 µm), kann fehlen (vielleicht epizooisch) Ventral:
Dasydytes (D. ) ornatus Dorsal: terminal 2 dick Stacheln (78-85 µm), distal mit Doppelspitze, an der proximalen Nebenspituze stark abgewinkelt; 8 Paare Stachelbündel: Kopf 1x; Hals 2x; 6 Paar Rumpf 3-5 x; längste stacheln 84-86 µm; am Kopf 3 weiter Stacheln, mittlere ohne Nebenspitze Ventral: 5-6 Paar Wimpernbüschel
Dorsale Schuppen: Hinten: 2 stumpfe Lappen mit kurzem Stachel (17-20 µm) mit Doppel- und Nebenspitze; auf einer Seite 12, auf anderer Seite nur 3 z.T. riesige Stacheln mit Doppel- und Nebenspitze; 3 dünne Stacheln am Hinterende (50µm)
Ventrale Schuppen: 4-5 Paar Zilienbüschel, sonst nackt
Ökologie: über Faulschlamm, Lemnatümpel
Ähnliche Arten: durch asymmetrische Stacheln einzigartig
Dorsale Schuppen: Auf den Hinterlappen ein Paar langer, auf Schuppenrudimenten sitzende Stacheln (63-68 µm; 1/4 Körperlänge); 2x7 Stachelbündel mit proximaler Nebenspitze; Kopf 2x mit 1 bzw 3 Stacheln; Rumpf 5x mit 4-7 Stacheln; Stachelanzahl und -länge (60.76µm) variiert
Ventrale Schuppen: 5 Paar Wimpernbüschel
Ökologie: semiplanktisch über Schlamm
Ähnliche Arten: evtl. synonym mit
D. (D. ) ornatus
; Unterschied Stacheln
Besonderheiten: sehr viele gleichmäßiger verteilte Stacheln
Dorsale Schuppen: Kopf zwei Ringe mit je 6 Stacheln mit einer Nebenspitze (10-15 µm); Körper 4 Paare Stachelgruppen mit je 5 Stacheln mit zwei Nebenspitzen (69-115 µm); Hinten zwei Paare Stacheln mit zwei Nebenspitzen (18µm); zwei Paar einfache Stacheln
Dorsale Schuppen: neun Paare Stachel-Gruppen; Stachelzahl von vorne: 1 - 1 - 2 - 5 - 5 - 3 - 1 - 1 (58-75 µm); erste beiden Gruppen einfach, sonst zwei Nebenspitzen; auf Schuppenrudimenten; Schlägerartige unpaarige Struktur am Hinterende (59x17 µm), kann fehlen (vielleicht epizooisch)
Ökologie: zwischen Wurzeln; Brasilien
Ähnliche Arten: D. (D. ) ornatus
hat weniger Stachelgruppen und keinen “Schläger”
Dorsale Schuppen: terminal 2 dick Stacheln (78-85 µm), distal mit Doppelspitze, an der proximalen Nebenspituze stark abgewinkelt; 8 Paare Stachelbündel: Kopf 1x; Hals 2x; 6 Paar Rumpf 3-5 x; längste stacheln 84-86 µm; am Kopf 3 weiter Stacheln, mittlere ohne Nebenspitze
1-3 Paar ventrolateral antspringende, kräftige Spungstacheln
hintere Sprungstacheln überragen den Rumpf um bis zu einer Körperlänge
weitere dorsale Sprungstacheln
keine Nebenspritzen
ventrale Wimpern in Reihen, keine Büschel
springende Bewegungen zur Flucht durch Sprungstachel möglich
Arten
6 Arten:
Haltidytes crassus Dorsal: keine Schuppen; 5 Paar sich nicht überkreuzende elastische, den Körper überragende Stacheln; je zwei Stacheln beginnen ventral Ventral: keine Schuppen; 3 Paar lange Sprungstacheln (100 - 240 mm); hintere überragen den Körper um eine Körperlänge und überkreuzen sich
Haltidytes festinans Dorsal: keine Schuppen; dorsolateral je 3 Gruppen zu 3 + 2 + 2 Sprungstacheln, die auch in Ruhelage abstehen und den Körper etwas überragen Ventral: keine Schuppen; ein Paar langer Sprungstacheln (140-144 µm), die den Körper um 3/4 seiner Länge überragen und sich überkreuzen
Haltidytes saltitans Dorsal: keine Schuppen; zwei lange, gerade caudale Sprungstacheln (70-75 µm); 4 Paar dorsolaterale sich überkreuzende Sprungsstacheln Ventral: keine Schuppen; ein Paar langer, sich überkreuzender Sprungstacheln (80-82 µm)
Dorsale Schuppen: keine Schuppen; 5 Paar sich nicht überkreuzende elastische, den Körper überragende Stacheln; je zwei Stacheln beginnen ventral
Ventrale Schuppen: keine Schuppen; 3 Paar lange Sprungstacheln (100 - 240 mm); hintere überragen den Körper um eine Körperlänge und überkreuzen sich
Ökologie: über Schlamm, früher häufig, imzwischen selten
Ähnliche Arten: H. festinans
: geringere Anzahl von Sprungstacheln
Besonderheiten: Setae stehen auf deutlichen Papillen
Haltidytes crassus war zur Zeit seiner Erstbeschreibung eine relativ häufige Art. Seither ist - wohl durch den Einfluss der industriellen Landwirtschaft - dieser hübsche Gastrotrich sehr selten geworden. Das Hauptmerkmal von H. crassus sind die fünf dorsalen und drei ventralen “Sprungsstacheln”, mit denen sich das Tier bei Bedrohung aus der Gefahrenzone katapultiert.
Dorsal; ds: dorsale Stacheln (5 Paar)
Auffällig ist das lange, caudale und haarförmige letzte ventrale Stachelpaar, das den Körper weit überragt und sich in Ruhestellung überkreuzt.
Zur schwimmenden Fortbewegung dienen drei Zilienringe am Kopf, während die für Gastrotrichen typischen ventralen Wimpernbänder zum Gleiten über das Substrat verwendet werden.
Ventral; cr: 3 Zilienringe zur Fortbewegung; js: Sprungstacheln; cs: lange Caudalstacheln; se: Seta auf Papillen
Die hinteren Tasthaare stehen - wie bei allen Mitglieder der Gattung Hatidytes - auf deutlichen Papillen.
Median; br: “Gehirn”; ph: Pharynx; oo: reife Eizelle; in: “Darm”; mf: Muskelfasern, die die Sprungstacheln bewegen
Die innere Anatomie wiest einige Besonderheiten auf. So sind deutliche Muskelfasern zu sehen, die an den Ansatzstellen der Sprungstacheln ankoppeln. Klebedrüsen und die zugehörigen Kleberöhrchen sind bei diese Gattung nicht vorhanden.
Die typische Bewegungsart und das Verhalten der Tiere ist in dem folgendem YouTube-Video gut zu sehen:
Dorsale Schuppen: 22-27 Reihen mit je 25-40 dünne, elliptische Kielschuppen (2 µm lang) ohne terminalem Stachel; Zehen unbeschuppt
Ventrale Schuppen: zwei kleine, gekielt Terminalplatten, sonst nackt
Ökologie: sehr weit verbreitet
Ähnliche Arten: H. ocellatum
: größere, weniger Schuppen; beschuppte Zehen und vZwf
Besonderheiten: meist relativ große Pseudozellen
Heterolepidoderma macrops ist eine der häufigsten und verbreitetsten Heterolepidoderma-Arten und wurde früher sicherlich oft mit
H. ocellatum
verwechselt, von der er sich - abgesehen von einigen schwer zu sehenden Einzelheiten - vor allem durch die größere Schuppenzahl unterscheidet:
Bei den meisten Tieren sind die an Augen erinnernde Pseudozellen sehr auffällig, die aber keine sensorische Funktion besitzen. Die hinteren Sinneshaare sitzen auf kleinen, dreieckigen “Sinnesschuppen”.
Dorsal mit Fokus auf die dorsalen terminalen Schuppen (dts)
Die Zehen der Tiere sind unbeschuppt. Das Schuppenkleid endet mit einem Paar relativ großer Kielschuppen an der Zehenbasis.
Querschnitt
Im Schnitt erkennt man, dass die kleinen Schuppen - im Gegensatz zu
H. ocellatum
- keinen distalen Fortsatz tragen. Der Pharynx ist an beiden Seiten leicht geschwollen.
Ventrale Ansicht mit Terminalplatten (tp)
Ventral trägt das Tier lediglich ein Paar kleiner, ovaler Terminalplatten. Der Rest des ventralen Zwischenfeldes ist - bis auf einige sehr dünne und schwer zu sehende ovale Schüppchen - unbeschuppt.
Die Haftröhrchen sind terminal verdickt.
Färbung der Zellkerne mit Methylgrün; B C D: Kerne des Gehirns; A: Übersicht
Färbt man H. macrops mit einem selektiven Kernfarbstoff wie Methylgrün, werden lediglich die Zellkerne der Tiere gefärbt. Dadurch erhält man einen guten Überblick über die Struktur des Gehirns. Deutlich ist in der Teilabbildung B der kernfreie Bereich dorsal des Pharynx zu erkennen, der bei (Zelinka, 1889)und (Remane, 1936)als “dorsale Kommissur”, also eine kernfreie Verbindung der beiden Gehirnhälften aus Nervenfasern, bezeichnet wird und die auch bei neueren Untersuchungen bestätigt wird (Schmidt-Rhaesa und Holger Rothe, 2014). Senkt man den Fokus auf die Mittelebene des Pharynx (Teilabbildung C) erkennt man den aus wenigen Zellen bestehenden Pharynx, der beidseitig von Gehirnzellen umgeben ist. Ventral ist keine Verbindung der beiden Gehirnhälften unterhalb des Pharynx zu erkennen. Das Gehirn umschließt also nicht den Pharynx vollständig und grenzt die Tiere zu den Cycloneuralia ab. Die bereits ungefärbt auffälligen Pseudozellen erweisen sich als mit Methylgrün färbbar - eine starkes Indiz dafür, dass es sich bei diesen Strukturen um die prominenten Zellkerne der sensorischen Nervenzellen der Tasthaare handelt.
Schuppenpräparation mit Eosin
Die Schuppenform der kleinen dorsalen Kielschuppen von H. macrops sind im Schuppenverbund am lebenden Tier nicht genau zu beurteilen. Eine genauere Schuppenpräparation zeigt eindeutig die ovale Form der Schuppen, die - im Gegensatz zu
H. ocellatum
- keine distale Spitze tragen.
Nach einer Phase mit parthenogenetische Fortpflanzung treten die Tiere in die sogenannte “Postpathenogenetische Phase” ein. In diesem Lebensabschnitt bilden sich die Tiere zu Zwitter um und entwickeln gleichzeitig Eier und Sperma, das in Bündeln ventral seitlich des Darms erzeugt wird - sie bilden sich zu echten Zwitter um.
Bei H. macrops sind die Spermatoiden spindelförmig. Gleichzeitig wird ein zweiteiliges, sogenanntes X-Organ mit unbekannter Funktion gebildet. Hier besteht das X-Organ aus zwei kugelförmigen, sekretgefüllten Zysten, die mit einem Steg (oder Verbindungsgang) verbunden sind.
Dorsale Schuppen: 25 Reihen mit je 40-80 länglichen engstehenden Kielschuppen (3 x4 µm, maximal 7 µm)
Ventrale Schuppen: 6-11 Reihen mit je 18-30 kleinen Kielschuppen (1,5 - 3 µm)
Ökologie: sehr häufig, auf Schlamm
Besonderheiten: Magenring, steht manchmal still
Heterolepidoderma majus ist ein sehr weit verbreiteter, schlanker und eleganter Bauchhärling.
H. majus: dorsale Beschuppung ( Ole Riemann)
Eines der beobachteten Tiere war in seiner postparthenogenetischen (Zwitter-)Phase und zeigte neben dem auffälligen, zweiteiligen X-Organ am Hinterende auch ein Sperma-Bündel:
H. majus: x-Organ (xO) und Spermienbündel (Sp) ( Ole Riemann)
Die Beschuppung an der Zehenbasis ist arttypisch. Die Sinneshaare entspringen aus Spezialschuppen:
H. majus: Beschuppung der Zehenbasis ( Ole Riemann)
Das ventrale Zwischenfeld ist mit kleinen, länglichen Kielschuppen besetzt und trägt zwei terminale Kielplatten:
H. majus: Beschuppung des ventralen Zwischenfeldes
Ein weiteres Merkmal, das die Art unverkennbar macht, ist der abgegrenzte “Magenring” am Vorderdarm:
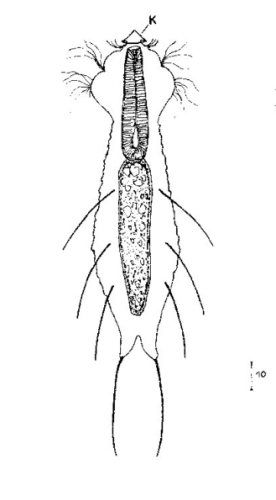
Ichthydium (F. ) tanytrichum Dorsal: nur dreieckige Basischuppen der terminalen Setolae Ventral: nackt bis auf zwei rechteckige, ungekielte Terminalplatten; Wimpernreihen am Rumpf in 4 Paar Wimpernbüschel aufgelöst; Wimpernbänder am Kopf vereinigt
Dorsale Schuppen: keine, hintere Setula auf Papille
Ventrale Schuppen: keine
Ökologie: Moor, auf Schlamm
Ähnliche Arten: Ichthydium monolobum wurde in diese Art aufgenommen
Besonderheiten: Sehr charakteristische Furca
Die Art I. (F.) forficula wurde vor einiger Zeit mit der etwas kleiner Art I. monolobum zu einer einzigen Art vereint. Ein Haupmerkmal dieser Art sind die langen, gebogenen und dünnen Kleberöhrchen, die den Tieren ein “pinzettenartiges” Aussehen verleihen:
I. forficula: dorsal mit pinzettenartigen Kleberöhrchen
Auf der Bauchseite trägt das Tier zwei deutliche Terminalplatten.
I. forficula: ventrale Terminalplatten
Die Wimpernbänder sind in einzelne Büschel mit extrem langen Wimpern aufgelöst.
I. forficula: Wimpernbüschel
In der Seitenansicht erkennt man die enorme Länge der lokomotiven Wimpern:
Dorsale Schuppen: nur dreieckige Basischuppen der terminalen Setolae
Ventrale Schuppen: nackt bis auf zwei rechteckige, ungekielte Terminalplatten; Wimpernreihen am Rumpf in 4 Paar Wimpernbüschel aufgelöst; Wimpernbänder am Kopf vereinigt
Ökologie: In Sphagnum / Moor
Ähnliche Arten: I. (F. ) skandicum
: Zusätzliche dorsale Schuppen an Hinterende
Besonderheiten: Sehr lange dorsal sichtbare Wimpern (27µm); sehr selten, bisher nur wenige Exemplare in Italien und Schweden
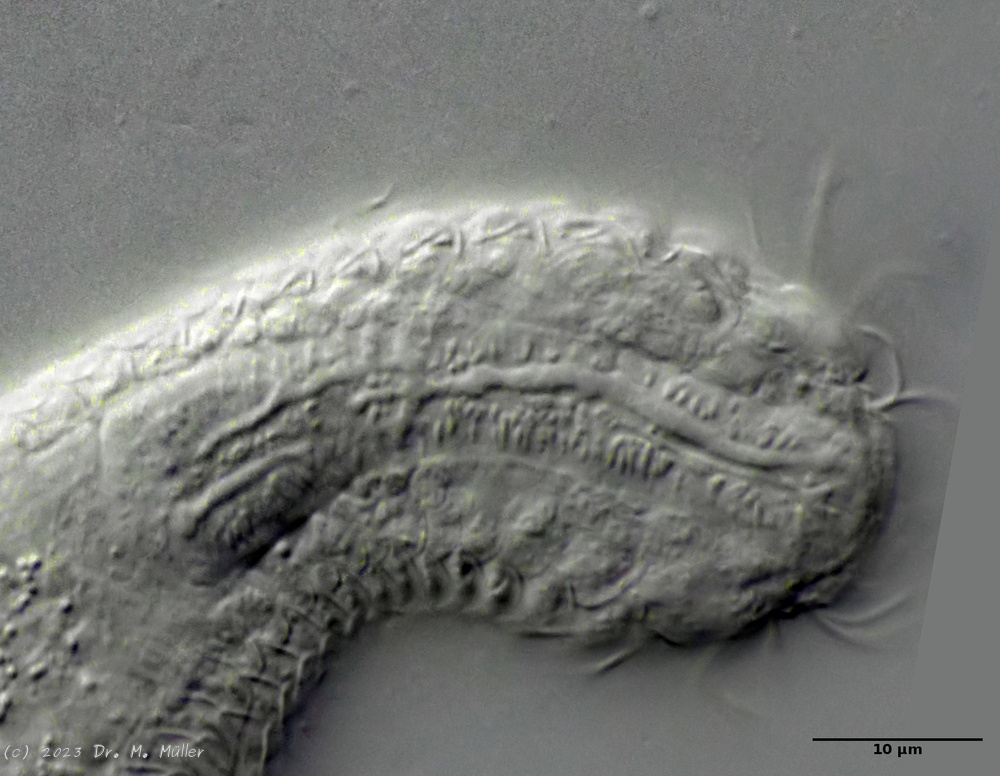
Ichthydium (Furficulichthys) tanytrichum ist ein sehr seltener Bauchhärling, der seit seiner Erstbeschreibung 1982 in Italien (Balsamo, 1982)nur noch aus Schweden - als ein Einzelexemplar - gemeldet wurde (Kånneby et al., 2009). Der hier gezeigt Fund aus dem Österreichischen Lauchseemoor ist das dritte weltweit bekannte Vorkommen dieser Art.
Mit einer Größe von ca. 130µm sind die Tiere der gefundenen Population etwa genauso groß wie die in Schweden gefundenen Tiere. Die italienischen Tiere der Erstbeschreibung waren signifikant kleiner (ca. 74µm). Bereits Kånneby äußerte den Verdacht, dass die Erstbeschreibung auf juvenilen Tieren fußt, da das von ihm gefundene erwachsene Tier mit 127µm deutlich größer war. Das würde auch die zur Erstbeschreibung etwas abweichenden Proportionen erklären.
Medianer Schnitt
Das Fehlen einer durchgehenden Beschuppung und die flexible Kutikula sind Kennzeichen der Gattung Ichthydium. Die typische zangenförmige Furka der Tiere stellt I. tanytrichum eindeutig in die Untergattung Furficulichthys, innerhalb der die Art durch Körperform und fehlende Beschuppung eindeutig festgelegt werden kann.
Dorsal: “Sinnesschuppen”, aus denen die Tasthaare entspringen
Weitere Artmerkmale sind die dreieckigen “Sinnesschuppen”, aus denen die hinteren Tasthaare entspringen. Das vordere Paar Tasthaare sitzt auf kleinen Papillen. Weiter Schuppen sind auf der Dorsalseite nicht vorhanden.
Ventral: In der Bauchregion sind die Wimpernbänder in Einzelbüschel aufgelöst
Auf der Bauchseite fallen die langen Wimpern auf, mit denen sich das Tier fortbewegt. Mit ca. 25µm Länge sind sie namensgebend für diese Art (“tanytrichum” = “langhaarig”). Typisch für diese Art ist, dass diese lokomotiven Cilien nicht - wie ansonsten bei Gastrotrichen üblich - in zwei durchgehenden Wimpernbändern organisiert sind, sondern in einzelnen Wimpernbüschel stehen. Terminal sind schwach die beiden großen, rechteckigen Terminalplatten zu erkennen.
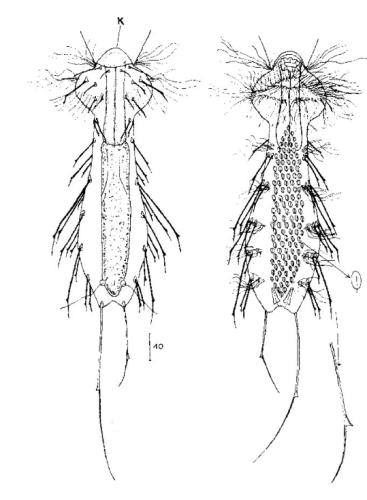
Lepidodermella minor Dorsal: 24 Reihen mit je 38 sehr kleinen, sich zum Til überlappenden kiel- und stachellosen Schuppen (1,2 x 2 µm) Ventral: Ventrales Zwischenfeld mit 6 Reihen 1,5 µm langen Kielschuppen und zwei ovale gekielte Terminalplatten (10µm)
Lepidodermella squamata Dorsal: 7-9 Reihen mit je 25-30 sehr unterschiedlich gestaltete, stachellose, stark überlappende Schuppen; Kopf- und Rumpfschuppen länglich oval mit dreieckigen / verbreitertem Vorderende (12 x 10 µm); Halsschuppen queroval, kurz (5 x 9 µm); Hypostomium: zwei Höcker Ventral: ventrales Zwischenfeld mit mehr als 20 Querspangen unter Pharynx und 3-5 Reihen viereckig abgerundeten, überlappenden Schuppen; 2 große Terminalplatten und einige kleine, runde Schuppen; Wimpernbänder vorne meist vereint
Eines der gefundenen Tiere hatte seine postpathenogenetische (Zwitter-) Phase erreicht und trug bereits ein X-Organ und Spermabündel:
L. minor chaetifer: oben dorsal mit den für Lepidodermella typischen glatten Schuppen; mitte im Querschnitt mit X-Organ und Spermabündel; unten ventrale Beschuppung mit ausgeprägten Terminalplatten ( Ole Riemann)
Im Sima-Moor wurde die etwas abweichende Unterart Lepidodermella minor chaetifer gefunden, die sich von der Art L. minor minor durch ein Paar Stacheln an der Zehenbasis unterscheidet:
L. minor chaetifer: Das Stachelpaar an der Zehenbasis ist charakteristisch für die Unterart chaetifer
Ein weiteres auffallendes Merkmal dieser Art ist der einzelne, asymmetrisch aus der Mundhöhle ragende Zahn:
Dorsale Schuppen: 7-9 Reihen mit je 25-30 sehr unterschiedlich gestaltete, stachellose, stark überlappende Schuppen; Kopf- und Rumpfschuppen länglich oval mit dreieckigen / verbreitertem Vorderende (12 x 10 µm); Halsschuppen queroval, kurz (5 x 9 µm); Hypostomium: zwei Höcker
Ventrale Schuppen: ventrales Zwischenfeld mit mehr als 20 Querspangen unter Pharynx und 3-5 Reihen viereckig abgerundeten, überlappenden Schuppen; 2 große Terminalplatten und einige kleine, runde Schuppen; Wimpernbänder vorne meist vereint
Ökologie: häufig, überall zu finden
Besonderheiten: durch rechteckige Schuppenplatten und große Terminalplatten gut abzugrenzen; sehr variable und gut untersuchte Art
Lepidodermella squamata ist einer der am besten untersuchten Gastrotrichen, da er in vielen Laboren weltweit in Kultur gehalten wird. Oft unterscheiden sich die “Kulturformen” aber bereits signifikant von den “Wildformen”, da die Tiere mit Ihrer enormen Generationenraten von etwa 50 pro Jahr rasch auf den Anpassungsdruck durch das Habitat reagieren können. L. squamata gilt deshalb als sehr variable Art. Die hier gezeigten Fotos stammen von Wildformen.
Dorsale Schuppen
Ventrale Schuppen; große Terminalplatten; rechteckige Spangen unter Hals
L. squamata ist eine der vielen Gastrotrichenarten, bei der sehr deutlich die Drüsengänge an der Oberfläche des Pharynx sichtbar ist. Diese Gänge verbinden einzelne Drüsenzellen in der Pharynxwand miteinander und leiten das Sekret der Drüsen in einen gemeinsamen Ausführungsgang zusammen, der sich in die Mundhöhle der Tiere ergießt:
Setopus bisetosus Dorsal: hintere Setolae auf kleinen Kielschuppen; Terminalstachel 45µm; Stacheln in 5 Gruppen zu je einem kurzem ventralen Stachel (10-15 µm) und einem längeren lateralen Stachel (ca. 40 µm); vordere und letzte Gruppe unpaar, also 8 Stacheln pro Seite Ventral: nackt; nur ein Paar Wimpernbüschel; Wimpernbüschel am Kopf typisch
Setopus primus Dorsal: keine Schuppen;Dorsolateral 3 Paar einzelne, leicht gebogene Stacheln (46µm); zwei einfache terminale Stacheln (40-54 µm) Ventral: keine Schuppen; 4 Wimpernbüschel in zwei Reihen
Setopus tongiorgii Dorsal: Terminalstacheln mit Nebenspitze auf dreieckigen Schuppenrudimenten; ungleich lang (27-30 µm; 43-55 µm); zahlreiche dorsolaterale Stacheln mit Nebenspitzen auf Schuppenrudimenten; pro Seite 21 Ventral: 5 Paar Wimpernbüschel; vZwf beschuppt (!) mit 8-9 Längsreihen winziger, ovaler Kielschuppen; zwei ca. dreieckige Terminalplatten mit Kiel (5 x 3,5 µm)
Dorsale Schuppen: hintere Setolae auf kleinen Kielschuppen; Terminalstachel 45µm; Stacheln in 5 Gruppen zu je einem kurzem ventralen Stachel (10-15 µm) und einem längeren lateralen Stachel (ca. 40 µm); vordere und letzte Gruppe unpaar, also 8 Stacheln pro Seite
Ventrale Schuppen: nackt; nur ein Paar Wimpernbüschel; Wimpernbüschel am Kopf typisch
Ökologie: pflanzenreiche Tümpel; über Schlamm; selten
Setopus bisetosus (Thomson, 1891) - eine früher häufige Art - ist durch das Fortschreiten der industriellen Landwirtschaft und der damit einhergehenden Trockenlegung sehr selten geworden. Deshalb freut es mich immer, ein Exemplar dieser Art zu finden.
freischwimmedes Exemplar
Leider konnte ich das Tier nicht vollständig still legen, so dass die Fotoqualität nicht optimal war.
Die semi-planktischen Setopus-Arten besitzen meist keine durchgehenden Wimpernbänder und bewegen sich mit einem (fast vollständig geschlossenen) Wimpernring am Kopf schwimmend. Zusätzlich sind ventral am Kopf einige Wimpernbüschel vorhanden. Am Körper selbst sind die Wimpernbänder - außer einigen vereinzelten Wimpern - auf ein Paar Wimpernbüschel am Hinterleib reduziert.
Im medianen Schnitt erkennt man zwei Paare gekielter Terminalplatten am Hinterende des Tieres. Diese Terminalplatten sind in der Literatur nicht erwähnt.
Lateral sind fünf Stachelpaare zu erkennen.
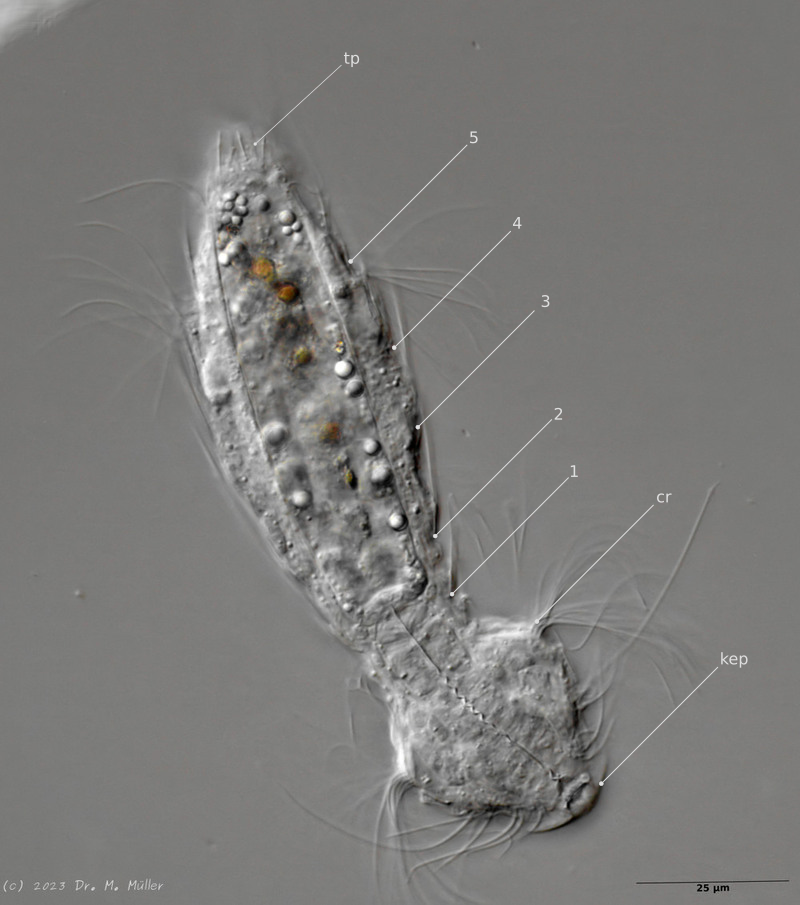
Mediander Schnitt mit Terminalplatten (tp) und Kopfschild (kep); 1..5: laterale Stacheln; Cilien-Ring (cr)
Typisch ist das breite und Starke Kopfschild (Kephalion) der Tiere.
Dorsale Schuppen: Terminalstacheln mit Nebenspitze auf dreieckigen Schuppenrudimenten; ungleich lang (27-30 µm; 43-55 µm); zahlreiche dorsolaterale Stacheln mit Nebenspitzen auf Schuppenrudimenten; pro Seite 21
Ventrale Schuppen: 5 Paar Wimpernbüschel; vZwf beschuppt (!) mit 8-9 Längsreihen winziger, ovaler Kielschuppen; zwei ca. dreieckige Terminalplatten mit Kiel (5 x 3,5 µm)
Ökologie: selten; Moorsee
Ähnliche Arten: S. chatticus
: keine Schuppen und weniger kürzere Stacheln
Besonderheiten: beschupptes vZwf
Setopus tongiorgii wurde bisher lediglich in Italien und einigen Standorten in Deutschland gefunden und ist - wie alle Setopus-Arten - relativ selten.
Überblick mit Fokus auf den Wimpernring am Kopf ( Dr. Stephan Krall)
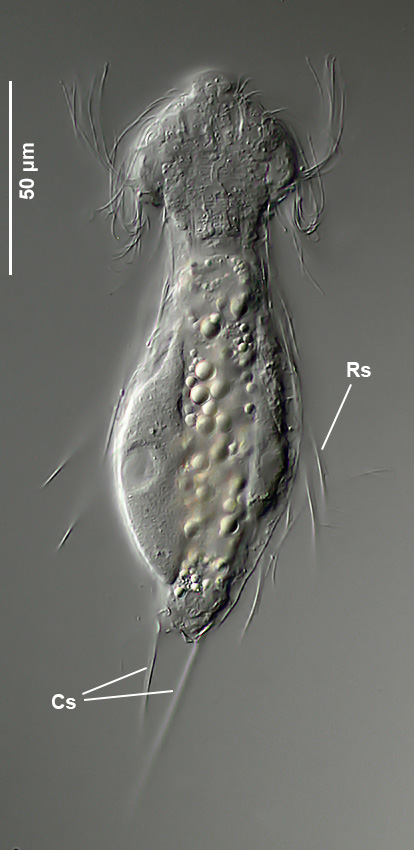
Querschnitt; Rs: Rumpfstacheln; Cs: caudale Stacheln ( Dr. Ole Riemann)
Im Querschnitt sind gut die seitlichen Stacheln mit ihrer Nebenspitze zu erkennen. Terminal stehen zwei ungleich lange caudale Stacheln, die ebenfalls eine Nebenspitze tragen.
Beschuppung des ventralen Zwischenfeldes ( Dr. Ole Riemann)
Ventral erkennt man die kleinen, rundlichen Schuppen und die gekielten Terminalplatten des ventralen Zwischenfeldes. Bisher wurden nur bei S. tongiorgii Schuppen in diesem Bereich gefunden. Diesem Merkmal kommt daher eine besondere Bedeutung bei der Artdiagnose zu.
Dieser Beitrag beschreibt lediglich die Anatomie von Süsswasser-Gastrotrichen. Die Anatomie von marinen Gastrotrichen kann beträchtlich abweichen!
Anmerkung
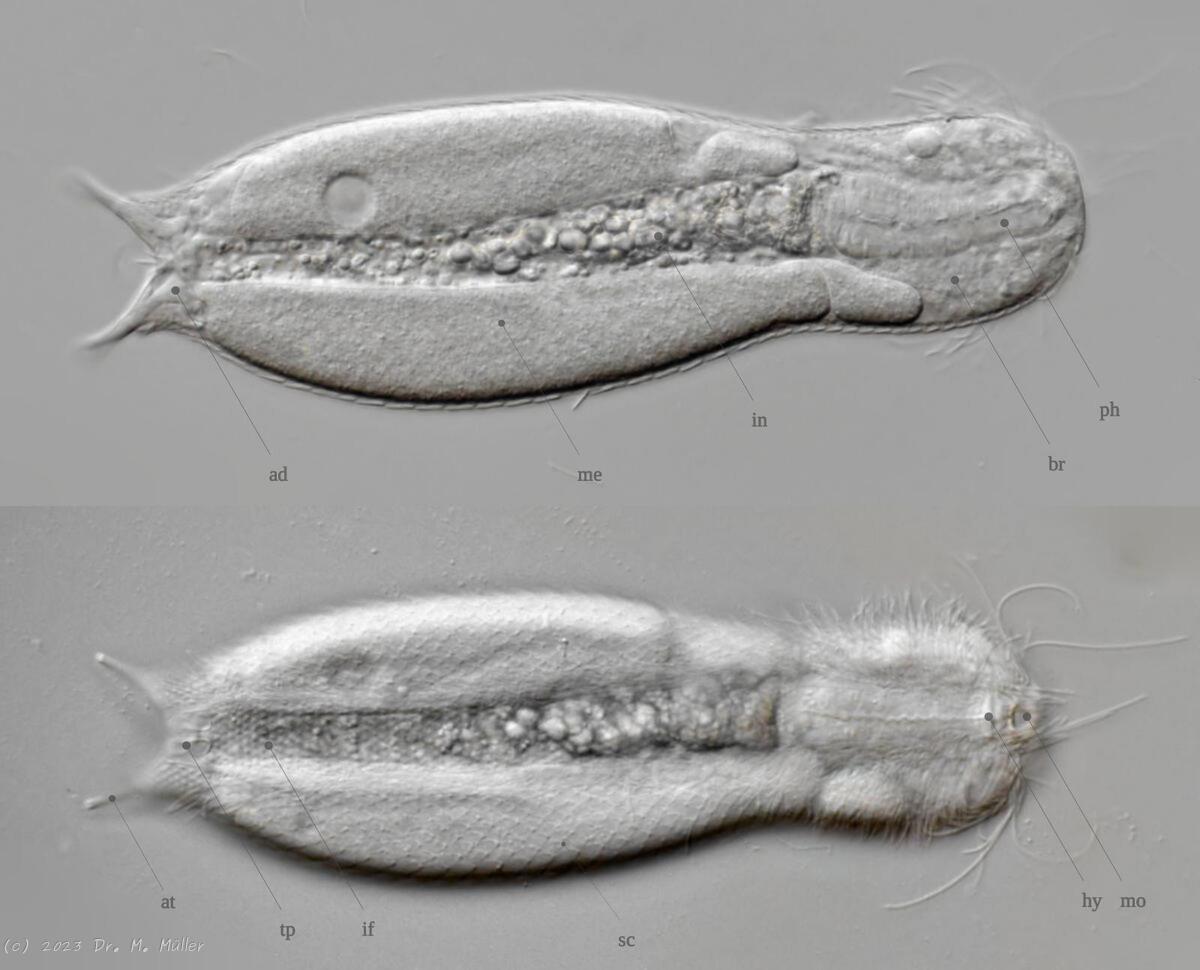
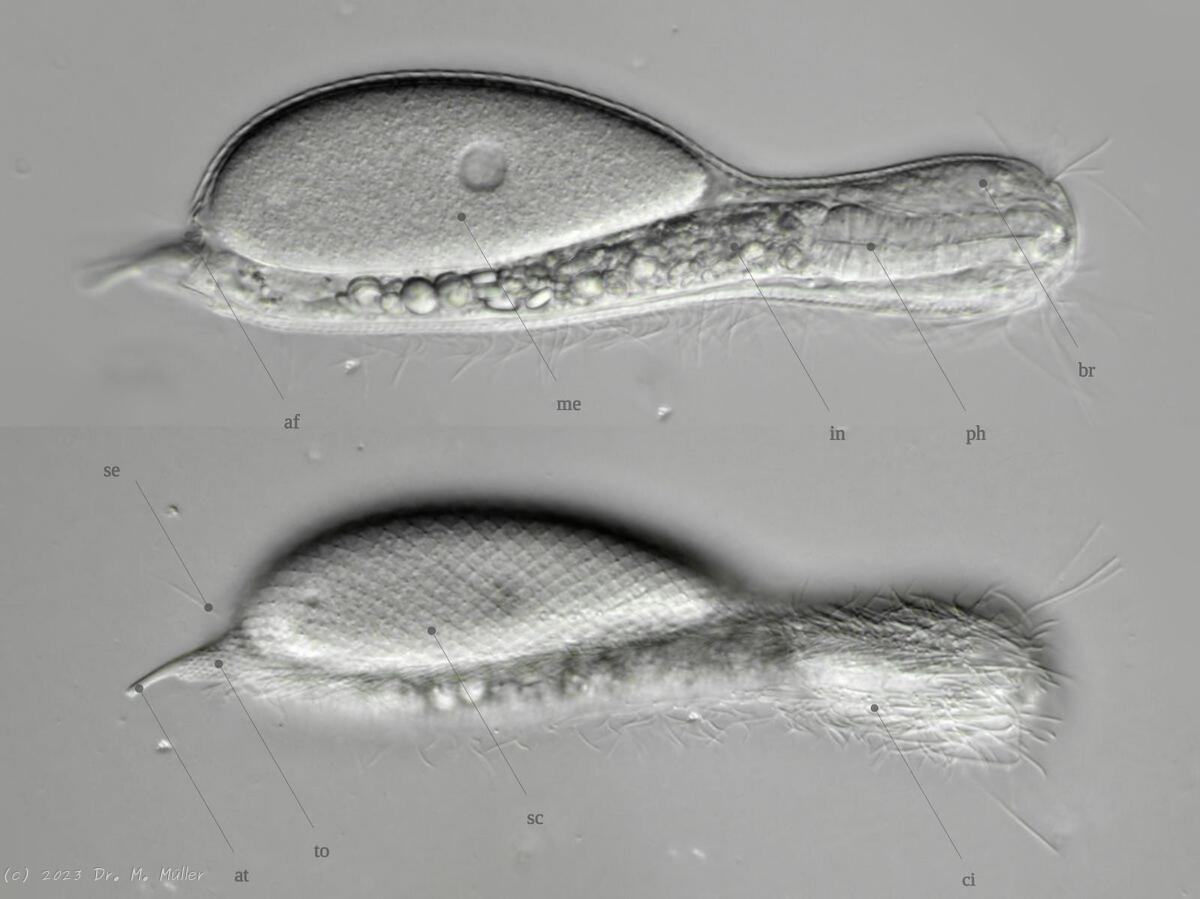
Bei einigen Gattungen und Arten ist die Anatomie in einigen Punkten abweichend zu dieser Darstellung!
Gastrotrichen gehören zu den kleinsten bekannten mehrzelligen Tiere, besitzen aber bereits viele der bei höher organisierten Lebewesen bekannten Organe. Die transparenten Tiere bieten deshalb einen einzigartigen Einblick in die grundlegenden Funktionen der Organsysteme für Nahrungsaufnahme und -verwertung, Fortpflanzung, Regulierung ddes Wasserhaushaltesund des Nervensystems bei einem zugleich sehr einfachen anatomischen Aufbau:
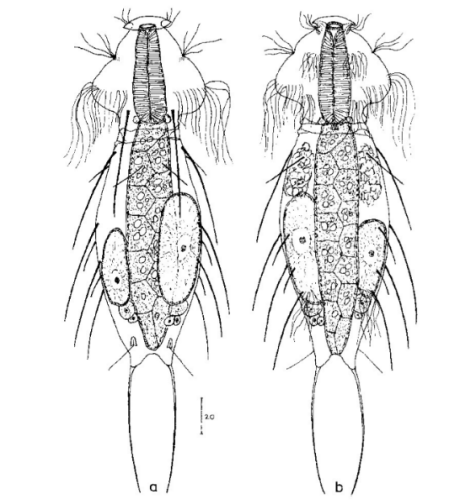
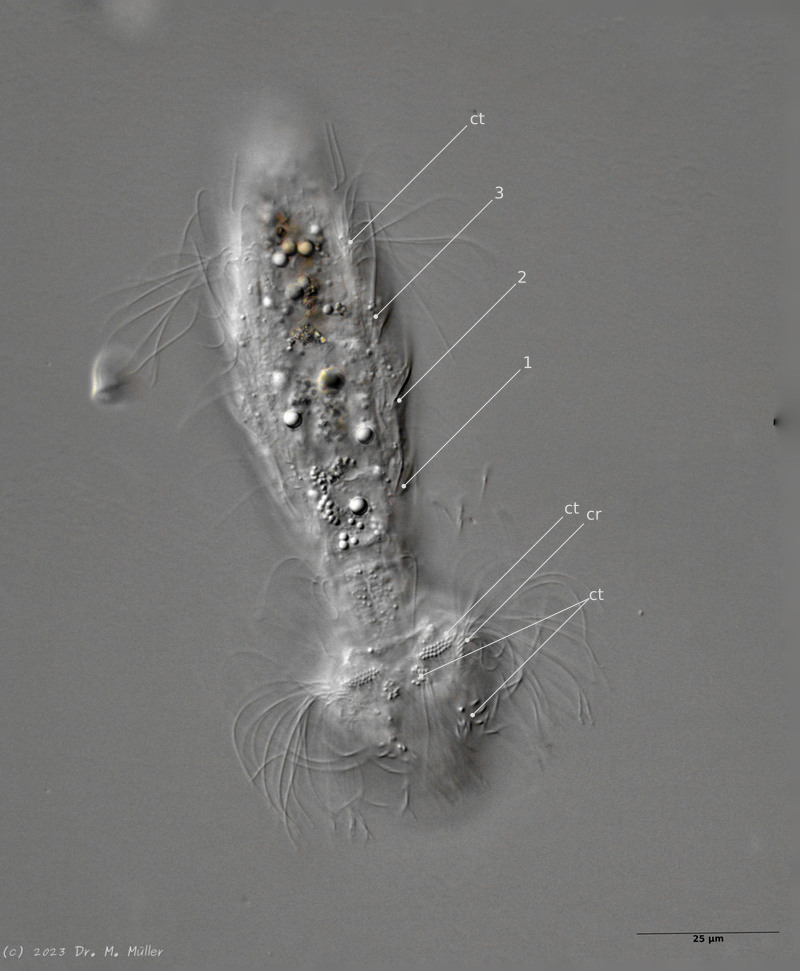
Die äußere Hülle der Gastrotrichen besteht aus einer dünnen und flexiblen Kutikula - einer Schicht aus anorganischem Material, die von den Hautzellen abgeschieden wird. Bei vielen Arten ist diese Kutikula von Schuppen (sc) bedeckt, deren Verteilung und Form sehr unterschiedlich und artspezifisch ist. Deshalb werden die Schuppen zur Artunterscheidung herangezogen. Zusätzlich zu den Schuppen sind noch weiter kutikulare Bildungen für Gastrotrichen typisch. Bei den meisten Arten (in der Unterfamilie Chaetonotoidae) endet der Körper in zwei Kleberöhrchen (at) auf zwei Zehenfortsätzen (to) in die die Gänge zweier Drüsen (ag) münden, die ein starkes Klebesekret produzieren. Mir diesem Sekret können sich die Gastrotrichen zum Schutz blitzschnell am Substrat festheften. Einige größere kutikulare Platten - z. B. das Hypostomion (hy) hinter dem Mund oder die Terminalplatten (tp) an der Bauchseite vor den Zehen - schützen empfindliche Bereiche der Tiere zusätzlich und sind von großer Bedeutung für die Artdiagnose.
An der Bauchseite besitzen die Gastrotrichen zwei namensgebende Bänder von Härchen (ci) (Gastrotrich=Bauchhärling), mit deren Hilfe sie über das Substrat gleiten oder im freien Wasser schwimmen. Zwischen den beiden Cilienbändern befindet sich das ventrale Zwischenfeld (if), das meist andere Schuppen als die Rückenseite trägt und das zur Artdiagnose von besonderem Wert ist.
Verdauungstrakt
Der Verdauungstrakt der Gastrotrichen ist relativ einfach aufgebaut. Der Mund der Tiere liegt am Vorderende der Bauchseite und besteht (meist) aus einer kurzen, runden, festen, kutukularisierten Röhre. Ihr Durchmesser ist meist sehr klein und kann nicht vergrößert werden (bei einigen Arten, z. B. den Arten der Untergattung Chaetonotus (Captochaetus) ist der Mund dehnbar und teilweise durch Lammellen verschließbar). Kurze, dünne Stacheln ragen aus der Mundöffnung und verschließen den Mund in Ruhestellung. Hinter dem Mund sitzt der kompliziert aufgebaute Pharynx (ph, “Schlund”), dem ich eine eigene Beschreibungsseite gewidmet habe. Der Pharynx besteht aus einer von Muskelfasern durchzogenen Röhre, deren Aufgabe es ist, den vom Substrat abgelösten Nahrungsbrei einzusaugen und die Nahrungspartikel aus der nahrhaften Suppe auszufiltern. Das überschüssige Wasser wird wider durch den Mund ausgeschieden und das so gewonnene Nahrungspellet weiter in den anschließenden Darm (in)befördert, in dem dei eigentliche Verdauung stattfindet. Der Darm besteht aus vier Säulen von Epithelzellen, die das Darmlumen umschließen. In diesem Hohlraum findet die Verdauung außerhalb der Zellen statt. Die Nährstoffen werden dann von den Wandzellen aufgenommen. Am Ende des Darms sitzt ein ringförmiger Schließmuskel (Sphincter), durch den die Verdauungsreste zurückgehalten werden, bis sie schließlich durch den meist auf der Rückseite sitzenden After (af) ausgeschieden werden.
Um den Pharynx auf der Rückenseite liegt das Gehirn (br) der Gastrotrichen. Die Bauchhärlinge besitzen also ein zentrales Nervensystem, von dem aus die entfernteren Organe mit feinen (lichtmikroskopisch praktisch nicht sichtbaren) Nervenfasern gesteuert werden. Die vorderen Tasthaare der Tiere entspringen direkt aus Nervenzellen im Gehirn während die Tasthaare (Setula) im “Nacken” und am Hinterende der Gastrotrichen eigene Nervenzellen besitzen, die über Nervenfasern mit dem Gehirn verbunden sind.
Fortpflanzung
Vom Fortpflanzungssystem sind meist nur die ungewöhnlich großen reifen Eizellen (me) zu bemerken. Gastrotrichen besitzen - relativ zu ihrer Größe - die größten Eier im Tierreich. Mit der komplexen Fortpflanzung beschäftigt sich ein eigener Abschnitt dieser Website.
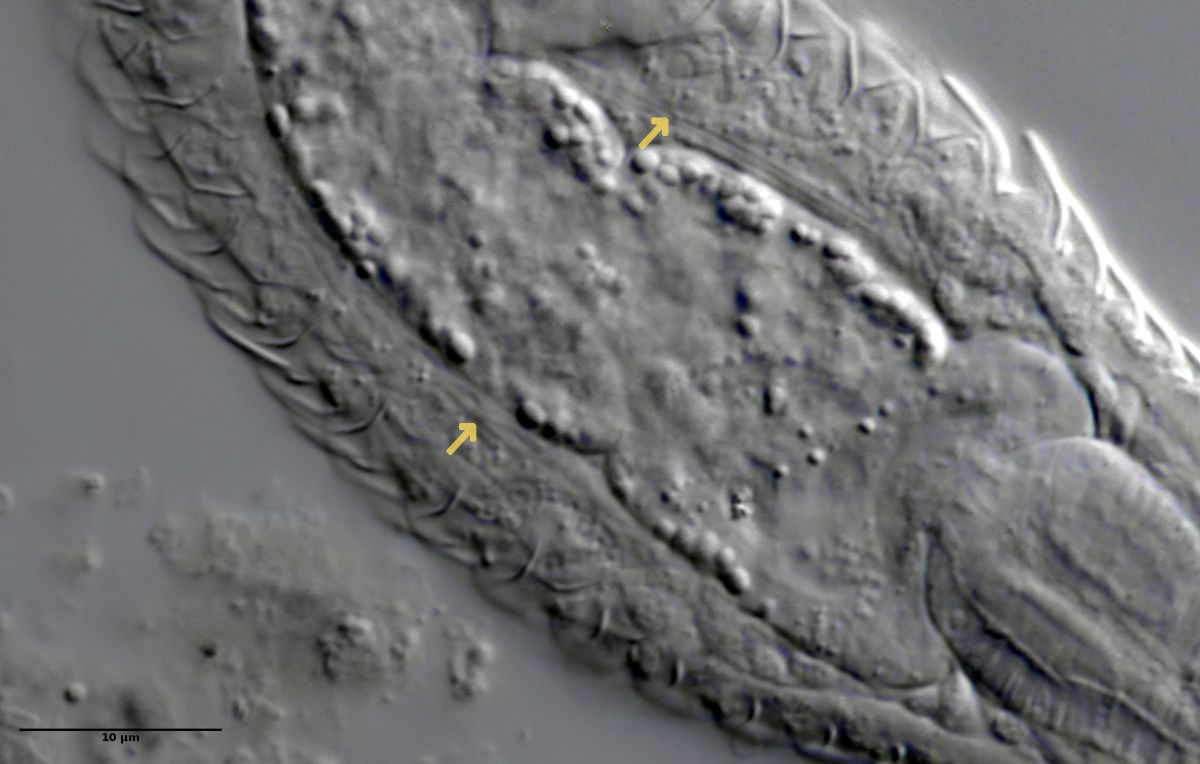
Prothonephridien
Abb. 3: Protonephridien eines typischen Gastrotrichen; Pfeile verweisen auf die beiden Wimpernkolben
Der Wasserhaushalt und Innendruck der Gastrotrichen wird durch zwei Vorläufer unserer Nieren, den Protonephriden reguliert. Diese Strukturen sich lichtmikroskopisch schwer zu erfassen und fallen oft nur durch einen flimmernden Bereich beidseitig des Darms der Tiere auf. Eine Protonephridie besteht aus einer einzelnen Zelle, aus der zwei lange, ständig schlagende Wimpern entspringen. Diese Wimpern befinden sich innerhalb eines kutikularisiertem dünnen Rohr, das zusammen mit den antreibenden Wimpern als “Wasserpumpe” arbeitet und “Wimpernkolben” genannt wird. Diese Röhre ist in Abb. 3, oberer Pfeil, gut zu erkennen. Beim unteren Pfeil erkennt man die beiden ineinander verschlungenen Wimpern, die den Wasserstrom aufrechterhalten. Aus elektronenmikroskopischen Untersuchungen ist bekannt, dass das in den Wimpernkolben strömende Wasser eine Siebplatte passieren muss, die Partikel zurückhält. Am andern Ende des Wimpernkolbens schließt sich ein langer, vielfach verschlungener Ausführungsgang an, der letztendlich im ventralen Zwischenfeld in das Umgebungswasser mündet.
Balsamo, Maria. 1982. „Three New Gastrotrichs from a Tuscan-Emilian Apennine Lake“. Bolletino Di Zoologia 49 (3–4): 287–95. https://doi.org/10.1080/11250008209439402.
(Balsamo & Todaro, 1982)
Balsamo, M. and M. A. Todaro. 1995. Gastrotrichi del Trentino: le Viotte del Monte Bondone (Trento). Studi Trentini di Science Naturali – Acta Biologica, 70: 9–22.
(Balsamo, 1983)
Balsamo M. 1983. Gastrotrichi. Guide C.N.R. per il riconoscimento delle specie animali delle acque interne, 20.
(Brunson, 1948)
Brunson,R.B. 1948. Chaetonotus tachyneusticus, a new species of gastrotrich from Michigan. Trans. Am. Microsc. Soc. 67: 350-351.
(Cordero, 1918)
Cordero,E.H. 1918. Notes sur les Gastrotriches. Buenos Aires, Phys. Rev. Soc. Arg. Sci. Nat. 4: 241-244.
(Dujardin, 1841)
Dujardin,F. 1841. Histoire Naturelle des Zoophytes, Infusoires. Librairie Encyclopedique de Roret,Paris 18: 568-570.
(Ehrenberg, 1831)
Suchen!
(Greuter, 1917)
Greuter,A. 1917. Beitrage zur Systematik der Gastrotrichen in der Schweiz. Rev. Suisse Zool. 25: 35-76.
(Garraffoni, 2017)
Garraffoni A.R.S., Araujo T.Q., Lourenço A.P., Guidi L., Balsamo M. 2017. A new genus and new species of freshwater Chaetonotidae (Gastrotricha: Chaetonotida) from Brazil with phylogenetic position inferred from nuclear and mitochondrial DNA sequences. Systematics and Bodiversity 15: 49-62.
(Gessener, 1930)
Gessener 1930, Mikrosk. f. Naturfreunde 8: 102-103, f. 6
(Gosse, 1864)
Gosse,P.H. 1864. The natural history of the hairy-backed animalcules (Chaetonotidae). Ananl. Magaz. nat. hist., 2 (8): 197-203
(Gosse, 1851)
Gosse,P.H. 1851. A catalogue of Rotifera found in Britain, with a discription of five new Genera and thirty-two new species. The Intellect. Observer 5: 387-406.
(Grosso & Drahg, 1991)
Grosso,L.E. and Drahg,F. 1991. Gastrotricos dulceacuicolos de la provincia de Tucuman IV. Acta Zool. Lilloana 40: 1.
(Grünspan, 1908)
Grünspan,T. 1908. Beitraege zur Systematik der Gastrotrichen. Zool. Jahrb. (Syst. ) 26: 214-256
(Horlick, 1975)
Horlick,R. 1975. Dasydytes monile, a new species of gastrotrich from Illinois. Trans. Ill. State Acad. Sci. 68: 61-64.
(d'Hondt, 1978)
d’Hondt,J.L. 1978. Gastrotricha, In: J. Illies, ed. Limnofauna Europa, Auflage 2. In Limnofauna europaea. Gustav Fischer, Stuttgart. pp. 92-97.
(Kånneby et al., 2009)
Kånneby, Tobias, M. Antonio Todaro, und Ulf Jondelius. 2009. „One New Species and Records of Ichthydium Ehrenberg, 1830 (Gastrotricha: Chaetonotida) from Sweden with a Key to the Genus“. Zootaxa 2278 (1): 26–46. https://doi.org/10.11646/zootaxa.2278.1.2.
(Kånneby, 2013)
Kanneby, T. 2013. New species and records of freshwater Chaetonotus (Gastrotricha: Chaetonotidae) from Sweden. Zootaxa 3701: 551-588.
(Kieneke und Schmidt-Rhaesa, 2014)
Kieneke, A. & Schmidt-Rhaesa, A. 2014. Gastrotricha. In: Schmidt-Rhaesa, A. (ed.), Handbook of Zoology. Vol.3 Gastrotricha and Gnathifera, pp 1-134. De Gruyter, Berlin, Boston.
(Kieneke und Riemann, 2008)
Kieneke, Alexander, und Ole Riemann. 2008. „First Record of Rare Dasydytid Gastrotrich Setopus Tongiorgii (Balsamo, 1982) from Central Europe with Some Biological Data“. Zootaxa 1680 (1): 59. https://doi.org/10.11646/zootaxa.1680.1.4.
(Kieneke, Martínez Arbizu, und Riemann, 2008)
Kieneke, Alexander, Pedro Martínez Arbizu, und Ole Riemann. 2008. „Body Musculature of Stylochaeta Scirtetica Brunson, 1950 and Dasydytes (Setodytes) Tongiorgii (Balsamo, 1982) (Gastrotricha: Dasydytidae): A Functional Approach“. Zoologischer Anzeiger - A Journal of Comparative Zoology 247 (2): 147–58. https://doi.org/10.1016/j.jcz.2007.11.001.
(Kisielewski, 1979)
Kisielewski,J. 1979. New and insufficiently known freshwater Gastrotricha from Poland. Ann. Zool. (Warsaw) 34: 415-435.
(Kisielewski, 1981)
Kisielewski,J. 1981. Gastrotricha from raised and transitional peat bogs in Poland. Monogr. Fauny Pol. 11.
(Kisielewski, 1986)
Kisielewski,J. 1986. Freshwater Gastrotricha of Poland. VII. Gastrotricha of extremely eutrophicated water bodies. Fragm. Faun. 30: 267-295.
(Kisielewski, 1987)
Kisielewski,J. 1987. Two new interesting genera of Gastrotricha (Macrodasyida and Chaetonotida) from the Brazilian freshwater psammon. Hydrobiologia 153: 23-30.
(Kisielewski, 1991)
Kisielewski,J. 1991. Inland-water Gastrotricha from Brazil. Ann. Zool. (Warsaw) 43 Suplement 2: 1-168.
(Kisielewski, 1999)
Kisielewski,J. 1999. A preliminary study of the inland-water Gastrotricha of Israel. Isr. J. Zool. 45: 135-157.
(Kolicka & Kisielewski, 2016)
Kolicka M., Dabert M., Dabert J., Kanneby T., Kisielewski J. 2016. Bifidochaetus, a new Arctic genus of freshwater Chaetonotida (Gastrotricha) from Spitsbergen revealed by an integrative taxonomic approach. Invertebrate Systematics 30: 398-419.
(Marcolongo, 1912)
Marcolongo,I. 1912. Gastrotrichi del lago-stagno di Astroni. Ann. Mus. Zool. Napoli nov. ser. Suppl 1,n° 2.
(Marcolongo, 1914)
Marcolongo,I. 1914. I Gastrotrichi del lago-stagno craterico di Astroni. Atti Reale Accad. Sci. Fis. Mat. ser. 2a 15: 1-25.
(Martin, 1981)
Martin,L.V. 1981. Gastrotrichs found in Surrey. Microscopy 34: 286-300.
(Metschnikoff, 1865)
Metschnikoff, E. 1865. Ueber einige wenig bekannte niedere Thierformen. Z. Wiss. Zool. 15: 450-463.
(Preobrajenskaja, 1926)
Preobrajenskaja,E.N. 1926. Zur Vrbreitung der Gastrotrichen in den Gewässern der Umgebung zu Kossino. Arb. Biol. Stat. Kossino Moskva 4: 3-14.
(Remane, 1927)
Remane,A. 1927. Beitraege zur Systematik der Susswassergastrotrichen. Zool. Jahrb. (Syst. Oekol. Geogr. Tiere) 53: 269-320.
(Remane, 1929)
Remane,A. 1929. Gastrotricha , In: W. Kükenthal & T. Krumback, eds. Handbuch der Zoologie, Band 2, Liefrung 6. Walter de Gruyter, Berlin.
(Remane, 1936)
Remane,A. 1936. Gastrotricha, In: H.G. Bronns, ed. Klassen Ordnungen des Tierreichs, Band 4, Abteilung II, Buch I, Teil 2, Lieferungen 1-2. Akademie Verlagsgesellschaft, Berlin.
Saito,I. 1937. Neue und bekannte Gastrotrichen der Umgebung von Hiroshima (Japan). J. Sci. Hiroshima Univ. 5: 245-265.
(Schimkewitsch, 1886)
Schimkewitsch,W. 1886. Uber eine neue Spezies Ichthydium. Nachr. Gesell. Freunde Naturw. Moskau 50: 148-150.
(Schmidt-Rhaesa und Holger Rothe, 2014)
Schmidt-Rhaesa, Andreas, und Birgen Holger Rothe. 2014. „6 Brains in Gastrotricha and Cycloneuralia – a Comparison“. In Deep Metazoan Phylogeny: The Backbone of the Tree of Life, herausgegeben von J. Wolfgang Wägele und Thomas Bartolomaeus, 93–104. DE GRUYTER. https://doi.org/10.1515/9783110277524.93.
(Schwank, 1990)
Schwank, P. (1990) Gastrotricha. In: Schwoerbel, J. & Zwick, P. (eds), Süsswasserfauna von Mitteleuropa, Band 3.Gastrotricha und Nemertini. Gustav Fischer Verlag, Stuttgart, Jena, New York, 252 pp.
(Schwank, 2014)
Schwank P., Kanneby T. 2014. Contribution to the freshwater gastrotrich fauna of wetland areas of southwestern Ontario (Canada) with redescriptions of seven species and a check-list for North America. Zootaxa 3811: 463–490.
(Stokes, 1887)
Stokes,A.C. 1887. Observations sur les Chaetonotus. J. de Micrographie 11: 77-84.
(Stokes, 1888)
Stokes,A.C. 1888. Observations sur les Chaetonotus et les Dasydytes. J. Microrg. 12: 19; 49-22; 51.
(Sudzuki, 1971)
Sudzuki,M. 1971. The Gastrotricha of Japan which live in the capillary water of the interstitial system: II. Bull. Biogeogr. Soc. Japan 27: 37-41.
(Thompson, 1891)
Thompson, P.G. (1891) A new species of Dasydytes, order Gastrotricha. Science Gossip, 319, 160–162
(Visvesvara, 1964)
Visvesvara,G. 1964. On some Gastrotricha of the genus Chaetonotus from India. Ann. Mag. Nat. Hist. 7: 209-216.
Eines der Hauptprobleme bei der Präparation von Gastrotrichen ist, die Tiere in einer Wasserprobe aufzuspüren und auf einen Objektträger zu pipettieren. Da Gastrotrichen transparent sind, sind sie in einem Stereo-Mikroskop mit traditioneller Auflicht- oder Durchlichtbeleuchtung nahezu unsichtbar. Die Tiere werden erst bei einer Dunkelfeldbeleuchtung sichtbar.
Im professionellen Bereich wird eine Dunkelfeldbeleuchtung durch Kaltlichtleiter realisiert. Eine Alternative Möglichkeit bieten LED-Leuchten mit Schwanenhals, die die Probe seitlich beleuchten. Durch diese seitliche Beleuchtung gelangt nur das an den Objekten gestreute Licht in das Objektiv des Mikroskops. Der Hintergrund bleibt reinschwarz. Da die Lampen nicht an der Probe befestigt sind, ändert sich die Beleuchtungsstärke und der Schattenwurf abhängig von der Probenposition. Oft stellen die Lampen auch ein mechanisches Hindernis bei der Verschiebung des Probengefäßes dar.
Meist ist diese Art der Beleuchtung bei Stereo-Mikroskopen nicht verfügbar und muss nachgerüstet werden.
Um diese Nachteile zu minimieren, habe ich für diesen Zweck habe ich eine seitliche Dunkelfeldbeleuchtung entwickelt, die mit einfachen Mitteln und einem 3D-Drucker leicht realisiert und an jedes Stereo-Mikroskop angepasste werden kann.
Halter für LED-Streifen
Drehbarer Objektführer
Um Objektträger entsprechend dem DIK-Gradienten oder dem Bildrändern der Kamera auszurichten, ist es nötig, die Präparate auf dem Objekttisch zu drehen. Wenn man - wie ich - keinen Drehtisch zu Verfügung hat muss man sich was einfallen lassen…
Ich habe mir einen drehbaren Objektführer konstruiert, der problemlos mit einen einfachen 3D-Drucker gefertigt werden kann. Dabei wurden folgende Eigenschaften berücksichtigt:
geringe Bauhöhe - dadurch sind Kollisionen mit den Objektiven ausgeschlossen
montiert an den Kreuztisch, um die Präparate weiterhin verschieben zu können
Präparat liegt direkt auf dem Objekttisch auf - keine Problem mit der Scharfstellung des Kondensors / Köhlern ist möglich
leichtes Einsetzen und Entnehmen des Objektträgers
abweichende Maße der Objektträger durch Federhalterung möglich
Einsetzen von den handelsüblichen Kunststoff-Petrischalen mit 7 cm Durchmesser möglich; zusätzliche kreisförmige Aussparung für die Verwendung der Deckel der Petrischalen (deren Bauhöhe etwas kleiner ist, so dass es zu keiner Kollision mit dem 10er Objektiv kommen kann)
Der Objektführer besteht aus zwei separaten Teile:
einem Drehrahmen der an den Kreuztisch mit den üblichen Befestigungsschrauben fixiert wird
einer Drehscheibe, die in den Rahmen gesetzt wird und in ihm gedreht werden kann. Um die Drehung zu erleichtern, liegt die Drehscheibe nur an sechs Punkten an dem Rahmen an. Die Drehscheibe ist mit sechs Griffen versehen, so dass sie leicht von allen Seiten bedient werden kann
Bild 1: Einzelteile des Objektführers; links Rahmen, der an den Kreuztisch montiert wird; rechts die Drehscheibe, die den Objektträger bzw. die Petrischale hält
Die beiden Teile werden ineinander gesteckt. Der Objektträger sitzt fest in der Aussparung und kann um 360 Grad gedreht und wie gewünscht ausgerichtet werden. Die runden Aussparungen sind ggf. für Petrischalen vorgesehen.
Bild 2: zusammengesteckter Objektführer
Am Mikroskop sieht das ganze dann so aus:
Bild 3: Drehbarer Objektführer am Mikroskop
In der Praxis hat sich diese Konstruktion bewährt. Natürlich ist der Drehtisch nicht zentrierbar - beim Drehen muss die Position des Objektes durch den Kreuztisch nachgeregelt werden. Die Objektträger sitzen fest genug, so dass es keinerlei “Schlupf” beim Verschieben gibt. Einige Objektträger-Marken scheinen aber eine etwas geringere Tiefe aufzuweisen, so dass man sich bei diesen wünscht, dass sie etwas straffer sitzen sollten. Will man solche Objektträger verwenden, muss man die Haltefedern etwas anpassen…
Die entsprechenden Druckfiles - inklusive den CAD-Files im OpenSCad-Format können bei Thingiverse heruntergeladen werden. Die Konstruktion ist auf den Objektisch meines Ortholux II angepasst. Für andere Mikroskope muss die Position bzw. der Durchmesser der Halteschrauben ggf. angepasst werden.
Ich möchte diesen praktischen Objektführer nicht mehr missen. Erst durch das Drehen des Objekts in den optimalen Winkel ist z.B. eine optimale Kontrastierung bei DIK möglich. Auch das Ausrichten an den Foto-Rändern erspart das ständige Drehen der Kamera.
Blitz
Die Vor- und Nachteile einer Mikroblitz-Einrichtung am Mikroskop wurde bereits in mehreren Beiträgen eingehend beleuchtet. Für mich als “Tümpler” war ausschlaggebend, dass ein Mikroblitz
es erlaubt, mit minimaler Lichtstärke (des Pilotlichtes) zu beobachten. Leider gibt es viele Organismen, die die oft hohen Lichtstärken, die bei DIK nötig sind, nicht gut vertragen und in Panik dem Licht ausweichen oder gar die Beleuchtung nicht überleben.
einen konstanten, von der Lichtstärke des Pilotlichts unabhängigen Weißabgleichs der Fotos gewährleistet.
kurze Belichtungszeiten unabhängig vom Pilotlicht ermöglicht. Will man mit der “normalen” LED-Beleuchtung kurze Belichtungszeiten realisieren, muss man vor dem Auslösen der Kamera typischerweise auf maximale Lichtstärke stellen, die aber für die Beobachtung unangenehm ist. Nur so erreicht man dann Belichtungszeiten kleiner als 1/500 s, die für ein “Einfrieren” der Bewegungsunschärfe nötig ist.
Bewegungsunschärfen mit den typischen Brenndauern von kleiner als 1/2000 bei ISO 100 minimiert. Mit LED-Beleuchtung erreiche ich bei 100er DIK bei ISO 400 typischerweise nur 1/200 s.
Deshalb habe ich mir Blitzeinrichtung für mein Ortholux II mit meinem 3D-Drucker konstruiert!
Bei einem Mikroblitz hat man prinzipiell die recht einfache Möglichkeit, den Blitz seitlich unter dem Kondensor einzuspiegeln. Ein solches Setup ist aber eher provisorisch, da beim “Normalbetrieb” Blitz und Strahlteiler unter dem Objekttisch stören und die ergonomische Bedienung einschränken. Für einen dauerhaft angebauten Mikroblitz an einem Mikroskop mit separaten Lampenhaus haben sich die “Stahlschmidt’schen Blitzwürfel” bewährt, die zwischen Lampenhaus und Stativ eingefügt werden und hier den Blitz in den “normalen” Strahlengang einspiegeln. So ist auch eine Blitzbeleuchtung unter “Köhlerbedingung” möglich, da die bestehende Leuchtfeldblende auch für den Blitz genutzt wird. Also musste es eine ähnliche Blitzwürfel-Lösung werden, bei dem die nötigen mechanischen Komponenten mit einem 3D-Drucker erstellt werden sollten. Meine Lösung sieht so aus:
Die zentrale Komponente ist der Halter des Strahlteiler (4), der schwenkbar ausgeführt würde. Dadurch ist es möglich, den Strahlteiler aus dem Strahlengang zu entfernen und den Beleuchtungsstrahlengang auch ohne Beeinträchtigungen durch den Blitz zu betreiben. Um einen sicheren Halt des ausgeschwenkten Strahlteilers zu gewährleisten, wurde der Schwenkgriff (10) mit einer Feder auch der Schwenkachse befestigt, so dass er gegen die Seitenwand drückt und in der oberen Stellung in die Aussparung einrastet. Zum Einschwenken des Strahlteiler muss der Griff nach Außen gezogen werden um eine freie Bewegung zu ermöglich. Lampen- und stativseitig wurden die Würfelwände (6 und 7) entsprechend den Maßen für das Lampenbajonett beim Ortholux II ausgeführt. Dadurch kann der Blitzwürfel zwischen Lampe und Stativ fest montiert werden und beeinträchtigt die Bedienung des Mikroskops nicht weiter. Die Lichtöffnung (2) am Deckel des Blitzwürfels wurde so an die Maße der Kollektorlinse angepasst, dass man sie einfach direkt einlegen kann. Bei einem Transport sollte deshalb die Kollektorlinse entfernt werden, da sie nur durch die Schwerkraft gehalten wird. Durch diese Konstruktion sitzt der Blitz direkt auf der Kollektorlinse und der Abstand zur Blitzröhre ist nahezu optimal. Durch die Fresnel-Platte des Blitzes können keine Störungen im Bild entstehen, da sie sehr nahe an der Kollektorlinse sitzt und deshalb nicht scharf abgebildet wird. Auf der Oberseite wird ein Halter (1) für den Blitz angeschraubt, der natürlich auf die Maße des verwendeten Blitzgeräts angepasst ist.
Zusammengebaut sieht das ganze dann so aus:
Gesamtansicht
Die Außenmaße (ohne Blitzhalter) des Blitzwürfels betragen 90mm * 100mm * 105mm - es handelt sich also eigentlich um einen “Blitzquader”. Diese Maße sind durch die Abmessungen der Zukaufteile (Strahlteiler, Kollektorlinse, Blitz) und die Vorgaben des Lampenbajonetts bedingt. In der Realität am Mikroskop sieht die Mikroblitzeinrichtung dann so aus:
Blitz am Ortholux II
Als Blitz verwende ich einen Godox TT685II für Canon-Kameras mit drahtloser Fernsteuerung, der problemlos mit meiner EOS700D zusammen arbeitet. Der Blitz hat eine Leitzahl von 60 und damit genügend Power, um beim 100er DIK noch mit 1/8 der Maximalleistung richtig zu belichten. Ich betreibe den Blitz im TTL-Modus, der problemlos die Blitzstärke an Motiv / Objektiv anpasst. Lediglich eine Belichtungskorrektur von -1.7 EV musste eingestellt werden. Dies Belichtungskorrektur ist unabhängig vom verwendeten Objektiv, muss aber - wie bei normalen Fotos auch - manchmal ans Motiv angepasst werden. Um den Blitz mit einem Netzteil zu betreiben, habe ich die Batterien mit Batteriedummies ersetzt, an die ein 6V Netzteil angeschlossen sind. Für alle, die sich an einen Nachbau wagen wollen, habe ich die entsprechenden OpenSCAD-Dateien hier zum Download zusammengestellt. Eine Anpassung an die Geometrie anderer Strahteiler, Kollektorlinsen und Blitzgeräte ist problemlos möglich. Schwieriger wird die Anpassung an Lampenbajonette Mikroskop-Hersteller. Das ist zwar prinzipiell möglich, erfordert aber u. U. Änderungen der Strahlführung, da sich dann möglicherweise die Lage der Lichtachse ändert. Bei Problemen einfach nachfragen - alles ist irgendwie möglich…
Ich bin mit dieser Mikroblitz-Lösung sehr zufrieden und möchte sie nicht mehr missen. Leider fällt damit eine weitere Ausrede für meine Foto-Qualität weg - jetzt muss ich wohl doch lernen, wie man gute Fotos macht…
Deckglas-Kompressorium
Um lebende Gastrotrichen detailliert mikroskopisch zu beobachten, ist es nötig,
dies Schichtdicke zu minimieren, da nur so die optimalen optischen Bedingungen für Objektive mit großen Vergrößerungen gewährleistet werden können
die Tiere unbeweglich festzuhalten
die Austrocknung der Präparate soweit zu minimieren, dass eine bequeme Beobachtungsdauer erreicht werden kann
das Eindringen von Immersionsöl in das Präparat zu verhindern
Zu diesem Zweck habe ich das Deckglass-Kompressorum entwickelt, das diese Bedingungen sehr einfach und schnell erreichen lässt.
Zunächst ist es wie immer nötig, das Beobachtungsobjekt mit einer dünnen Pipette unter dem Stereo-Mikroskop zu isolieren und mit einem möglichst kleinen Wassertropfen auf eine gereinigten Objektträger aufzubringen. Dabei sollte darauf geachtet werden, dass der Wassertropfen so knapp bemessen wird, dass die Wasserschicht unter dem aufzubringendem Deckglas bei der gewünschten Schichtdicke die Ränder des Deckglases nicht erreicht. Hierzu ist etwas Erfahrung nötig…
Deckglas-Kompressorium: Vaselinestege in gelb
Etwas Vaseline wird auf einem Handballen verrieben. Mit dem oberen und unteren Rand einem rechteckiges Deckglas (ich verwende meist die Größe 18x18mm) streift man einen dünnen Steg Vaseline von dem Handballen ab. Die Vaseline dient als Abstandshalter une Randabschluss des Deckglases. Das Deckglas wird mit einem Rand schräg auf den Objektträger gesetzt - die Vaseline hält das Deckglas in dieser Stellung. Mit einem Holzstöckchen wird der abstehende Rand des Deckglases vorsichtig zum Objektträger gedrückt, bis es den Wassertropfen erreicht und auch mit der oberen Seite auf dem Objektträger anliegt. Der Wassertropfen füllt nun mit relativ großer Schichtdicke einen kleinen Teilbereich des Deckglases aus.
In diesem Zustand ist der im Wassertropfen gefangene Gastrotrich noch frei beweglich und kann bequem mit schwächeren Objektiven beobachtet werden. Durch die kleine Fläche des Wassertropfens ist das Auffinden des Objektes problemlos möglich.
Unter Mikroskopkontrolle kann man nun vorsichtig die Schichtdicke des Wassers durch Festdrücken der Deckglasränder gerade soweit verringern, dass der Gastrotrich nicht mehr beweglich ist und sanft durch das Deckglas auf den Objektträger gedrückt wird. Die beiden Vaseline-Ränder setzen dabei dem manuellen Druck genügend Widerstand entgegen um die Schichtdicke sehr kontrolliert verringern zu können.
Durch die beiden gegenüber liegenden Vaselineränder wird gewährleistet, dass
das Eintrocknen des Präparates stark verzögert wird, da eine Verdunstung des Wassers nur an den freiliegenden Deckglasrändern ohne Vaseline erfolgt und deshalb stark reduziert ist.
die eingestellt Schichtdicke sich beim Austrocknen des Präparates nahezu unverändert bleibt, da der Wassertropfen nur an den Rändern verdunstet und sich lediglich sein Durchmesser ändert.
ein Eindringen von Immersionsöl an den abgedichteten Deckglaskanten unmöglich ist, an den freiliegenden meist keinen Kontakt mit dem Probenwasser besteht.
ein “Durchziehen” von Chemikalien zum Färben etc. wegen der gegenüberliegenden offenen Deckglaskanten problemlos möglich ist.
ein späteres manuelles Verringern der Schichtdicke bis zum Plattdrücken des Objektes (z. B. zur Schuppenanalyse jederzeit kontrolliert möglich ist
Mit etwas Erfahrung ist die Erstellung eine Deckglas-Kompressoriums problemlos und innerhalb von wenigen Sekunden möglich.
Ich verwende dies Präparationsart für hochauflösende Beobachtungen seit Jahren und möchte sie nicht mehr missen!
Doppeltes Deckglas
Gastrotrichen sind Tiere, bei denen sich die Ober- und Unterseite stark unterscheiden. Oft ist es nötig, sowohl die ventrale als auch die dorsale Seite genau zu beobachten. Wenn man eine Tier mit einem Deckglas-Kompressorium beobachtet, kann meist nur eine Seite exakt beobachtet werden. Die andere Seite ist nur zugänglich, wenn man durch das Tier hindurch fokussiert. Dies hat aber - gerade bei hoch auflösenden Objektiven - große optische Qualitätseinbußen zufolge. Man kann dann nur versuchen, das Tier durch Verschieben des Deckglases zu drehen. Leider hat diese Methode aber nur bei drehrunden Objekten eine Aussicht auf Erfolg und endet bei Gastrotrichen meist letal… :cry:
diagonales doppeltes Deckglas Vaselinesteg in gelb
Um eine beidseitige Beobachtung zu ermöglichen, kann man das Objekt zwischen zwei Deckgläser einschließen und bei Bedarf den gesamten Deckglasstapel auf dem Objektträger wenden. Dazu verwendet man zwei unterschiedlich große Deckgläser. Das größere Deckglas (z. B. 20x20 mm) heftet man mit einem kleinen Tropfen destilliertem Wassers auf einen Objektträger. Das Deckglas sollte diagonal ausgerichtet sein, um es bei Bedarf leicht an den gegenüberliegenden Ecken greifen zu können. Auf dieses Deckglas bringt man mit einer Pipette das zu beobachtende Objekt. ein zweites, kleineres Deckglas (18x18 mm) wird an zwei Rändern (wie im Beitrag Deckglas-Kompressorium beschrieben) mit einem Vaselinesteg versehen kontrolliert auf das untere Deckglas mit der Probe gedrückt und das Objekt dadurch zwischen den beiden Deckgläsern festgehalten.
Das Objekt kann nun mit der gewohnten optischen Qualität an der Oberseite beobachtet werden. Ist die Beobachtung von dieser Seite aus beendet, wird (wenn noch nicht erfolgt) ein Tropfen Immersionsöl auf das Deckglas gegeben und der Deckglasstapel an den überstehenden Ecken vom Objektträger gelöst. Der Stapel wird gedreht und mit der eingeölten Seite auf den Objektträger gelegt. Jetzt kann die Beobachtung von der nun oben liegenden, früheren Unterseite aus erfolgen. Durch die Verwendung unterschiedlicher Deckglasgrößen wird sichergestellt, dass man beim Drehen des Deckglasstapel nur ein Deckglas berührt und die Deckgläser nicht gegeneinander verschiebt.
Mikroaquarium
Unter Mikroaquarien verstehe ich Präparate in Objektträgergröße, die man direkt mit dem Mikroskop beobachten kann. Dabei sollte die optische Qualität der Mikroaquarien (speziell die Schichtdicke) gut genug sein, um jede mögliche Vergrößerung zu verwenden. Außerdem sollte die Lebensdauer der Organismen zumindest einige Wochen betragen, um das Verhalten bzw. die Entwicklung der Objekte studieren zu können. Nicht zuletzt muss das Verfahren einfach und schnell anwendbar sein - ein umständliches und mit einem hohen apparativen Aufwand verbundenes Verfahren ist in meinen Augen im Amateurbereich nicht praktikabel. Vor einigen Jahren bin ich im Internet auf eine Präparationsmethode gestoßen, die ich zu einem Langzeit-Mikroaquarium weiterentwickeln konnte. Ein solches Mikroaquarium ist folgendermaßen aufgebaut:
Aufbau eines Mikroaquariums
Die Lebensdauer eines “normalen”, temporären Tümpler-Präparates kann erhöht werden, wenn man das Verdunsten des Wasser effektiv verhindert. Dies kann erreicht werden, wenn man das Deckglas mit einem Vaseline-Rand wasserdicht versiegelt. Vaseline besitzt zu diesem Zweck eine Reihe von idealen Eigenschaften:
sie ist absolut wasserdicht
Vaseline ist luftdurchlässig (kennt man von Feuchtigkeitscremes auf Vaselinebasis)
wegen ihrer Konsistenz ist Vaseline leicht zu verarbeiten
sie ist in jedem Drogerie-Mark preiswert zu finden
Um das Deckglas mit einem Vaselinerand zu versehen, trägt man einen dünnen (!) Vaselinefilm auf seinen Handballen auf und streift mit jedem Deckglasrand einen dünnen Steg von der Haut ab. Das Deckglas kann dann direkt mit einem Rand auf den Objektträger (mit Probe) gesetzt werden. Von diesem Rand aus senkt man das Deckglas ab und drückt das Deckglas mit einem Holzstäbchen langsam auf die gewünschte Schichtdicke zurecht. Überschüssiges Wasser wird dadurch unter dem Deckglas heraus gedrückt und kann dann leicht entfernt werden. Um ein langlebiges Mikroaquararium zu erstellen, ist es nötig eine relativ große Wassermenge im Präparat einzuschließen. Ist dieser Wasserpuffer zu gering, würden die Organismen nach wenigen Tagen versterben. Deshalb verwendet man für ein Mikroaquarium einen Objektträger mit Hohlschliff. Um die zur Beobachtung nutzbare Fläche zu erhöhen, verwende ich ein möglichst großes Deckglas (z. B. 24mm x 32 mm oder 24mm x 60mm). Dadurch ist die Fläche außerhalb des Hohlschliffs - die Fläche mit der optimalen Schichtdicke - maximiert. Ein so erstelltes Mikroaquarium hat viele Vorteile:
Lebensdauer der Organismen bis zu 6 Wochen
große mechanische Stabilität:
durch das festsitzende Deckglas kann ist die Reinigung problemlos
senkrechte Aufbewahrung in normalen Präparateboxen (transparente Abdeckung!)
absolut stabile Verhältnisse auch für Langzeit - Videoaufnahmen
Einsatz von Immersionsöl unproblematisch: DG verrutscht nicht, Öl zieht nicht unter das Deckglas
Zusatzaufwand zur Erstellung nur wenige Handgriffe
Der Hauptvorteil ist aber in meinen Augen, dass sich die Organismen nach einer Eingewöhnungszeit von ca. einem Tag in den Mikroaquarien “natürlich” verhalten. Die typische große Unruhe, wie man sie aus den “normalen”, temporären Tümpler-Präparaten kennt, ist abgeklungen und die eingeschlossenen Tiere gehen zu ihrem “Tagesgeschäft” über. Die meisten Ciliaten weiden ruhig ihre Nahrung ab oder stehen sogar über Stunden an dem selben Platz und strudeln entspannt ihr Futter ein. Gastrotrichen beruhigen sich und tauchen nach einiger Zeit aus ihren Verstecken auf und weiden - optimal beobachtbar - die Unterseite des Deckglases ab. Um diese entspannten Verhältnisse in einem Mikroaquarium zu demonstrieren, möchte ich folgenden - nahezu unbearbeiteten Film - zeigen, der in mehreren Mikroaquarien einige Tage nach deren Erstellung aufgenommen wurde:
Zum Starten des Filmes bitte auf das Bild klicken!
Für mich ist diese Art der Präparation inzwischen die befriedigendste Art, die Wunderwelt im Tümpel zu beobachten. Natürlich ist es so nicht möglich, die letzten anatomischen Details der Organismen abzubilden. Dafür erhält man aber die Möglichkeit das Verhalten oder die Entwicklung der Organismen über einen langen Zeitraum und stressfrei zu verfolgen.
Schuppenpräparation
Bei Bauchhärlingen erfolgt die Arteinteilung traditionell über die Form der Schuppen. Daher ist die Artbestimmung oft nur möglich, wenn man die Schuppenform der Tiere genau beurteilen kann. Leider sind die Schuppen der Gastrotrichen oft nur schwer zu sehen. Selbst mit DIK - obwohl hier die schwache Doppelbrechung der Schuppen den Kontrast noch zusätzlich verstärkt - ist bei vielen Arten eine Lebendbeobachtung zur Artdiagnose nicht ausreichend. Deshalb möchte ich hier eine Möglichkeit zur Präparation der Schuppen und damit zur (meist) sicheren Artdiagnose vorstellen. Als Beispiel habe ich
Lepidochaetus zelinkai
gewählt. Durch das markante Erscheinungsbild dieses Gastrotrichen ist eine Schuppenanalyse zur Artbestimmung eigentlich nicht nötig, aber einige Schuppendetails werden in der Literatur kontrovers diskutiert, so dass ich mir mal selbst ein Bild machen wollte.
L. zelinkai
ist ein mittelgroßer Gastrotrich mit einem markanten Stachelkleid:
Selbst bei stärkerer Vergrößerung werden nicht alle Details der Beschuppung deutlich:
Bild 2:
L. zelinkai
: Schuppen des ventrales Zwischenfeldes
Um die Schuppen frei zu präparieren, muss das Tier mazeriert werden. Dazu legt man das Tier - am besten mit einem einfachen Vaseline-Kopressorium - mit dem Deckglas fest und zieht einen Tropen Eisessig unter das Deckglas. Den Eisessig habe ich vorab mit sehr wenig Eosin angefärbt - dadurch werden die Schuppen gleichzeitig bei der Mazeration gefärbt. Der Eisessig löst alle Weichteile des Tieres innerhalb weniger Minuten auf, während die Schuppen der Gastrotrichen säurebeständig sind und deshalb erhalten bleiben. Je nach Schuppengefüge verbleiben die Schuppen als zusammen hängende Hülle erhalten oder lösen sich voneinander. Bei
L. zelinkai
sind die Schuppen anscheinend nicht miteinander verzahnt, so dass die Schuppen sich verteilen:
Diese Art der Präparation ist recht einfach und erlaubt es, die Schuppenformen genau zu beurteilen. So fallen z.B. bei den obigen Bildern die Stege in der Schuppenfläche auf, die sich parallel zur Vorderkante erstrecken. In der Literatur werden diese “doppelten Ränder” der Schuppen meist als optisches Artefakt angesehen, das durch den zweifachen Schnitt der Fokusebene durch die gebogenen Schuppen erzeugt wird. Die obige Schuppenanalyse belegt eindeutig, dass diese auffälligen Stege wirklich existieren und eine Verdickung der Schuppen darstellen.
Webdesign
Der Plan, eine Website zu erstellen, die eine Hilfestellung bei der Bestimmung von Gastrotrichen bietet und meine eigenen Beobachtungen und Erkenntnisse systematisch zusammenfasst, ist bereits einige Jahre alt. Als ich mit der Arbeit hierzu begann, habe ich - wie wohl die meisten Amateure - meine Erfahrungen mit WordPress gesammelt.
WordPress ist ein wunderbares Content Management System, das sehr leicht zu bedienen ist - solange man sich an die Vorgaben und Vorgehensweisen hält, die WP vorgibt. Leider hatte ich für meine Website andere Vorstellungen als die Entwickler von WP und sehr bald musste ich feststellen, dass ich mehr Arbeit auf das Verbiegen von WP und das Anpassen des Designs aufwand, als für das Einbinden von Inhalten. Jede neue Art-Webseite musste in entsprechende Navigations-Menüs eingebunden werden, jede Änderung der Struktur der Website zog umfangreiche Änderungen in WP-Listen nach sich. Zusätzlich fand ich es schwierig, ein durchgehendes Webdesign für die Einzelseiten durchzuhalten, da das Design direkter Bestandteil des Inhalts der Webseiten war. Wenn man sich zu einem späteren Zeitpunkt für eine andere Darstellung der einzelnen Art-Seiten entschied, mussten sämtliche bestehenden Seiten angepasst werden. Irgendwann musste ich mir eingestehen, dass WP für meine Art zu arbeiten nicht geeignet ist und ich entweder meinen Workflow ändern oder auf die Vorteile von WP verzichten müsste.
Durch Zufall stieß ich auf den Webseitengenerator “HUGO”, der es erlaubt, lokal statische Webseiten zu generieren, die dann erst auf “Knopfdruck” auf einen einfachen Webserver kopiert werden - kein kompliziertes Datenbanksystem, keine Designvorgaben, keine proprietäre Plugins wie bei WP. Und das beste dabei: HUGO versucht sehr erfolgreich, Inhalt und Design streng voneinander zu trennen!
In HUGO wählt man ein “theme” als Anfangsdesign aus und passt es einmalig an seine Bedürfnisse an. Die Einzelseiten werden dann in einem einfachen Texteditor im “Markdown”-Format erstellt und meinen Designvorgaben entsprechend generiert und in die Website integriert. Dabei ist es möglich, Datenfelder in den Textdateien zu pflegen, deren Inhalte dann in die Darstellung der Webseite aufgenommen werden. So kann man z. B. für jede Gastrotrichenart ein einfaches, frei definierbares “Datenformular” auszufüllen, in dem allgemeine Größenangaben und Beschreibungen der Art eingegeben werden. Diese Angaben werden dann von HUGO verwendet um die entsprechende Webseite mit einem Standarddesign z. B. für einen “Art-Steckbrief” zu erzeugen. Daten und Design sind vollständig voneinander getrennt. Alle Webseiten verwenden ein konsistentes Design, das man zu jedem Zeitpunkt für alle Webseiten gleichzeitig verändern kann - ohne alle Einzelseiten anpassen zu müssen! Dazu kommen noch die Vorteile der Verwendung von statischen Webseiten, die eine unübertroffene Reaktionsgeschwindigkeit zu minimalen Kosten bei einer vollen Kontrolle über den Datenschutz bieten.
Für mich hat die Verwendung von HUGO die Erstellung und die Pflege meiner Website stark erleichtert und ich möchte mich herzlich bei den vielen ehrenamtlichen Entwicklern bedankten, die HUGO erstellt haben und kontinuierlich pflegen :heart: !
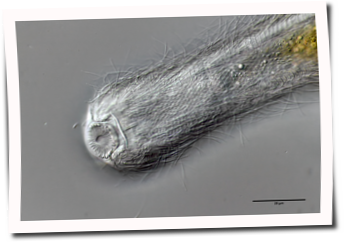

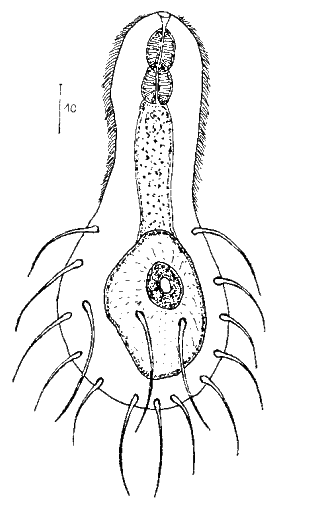
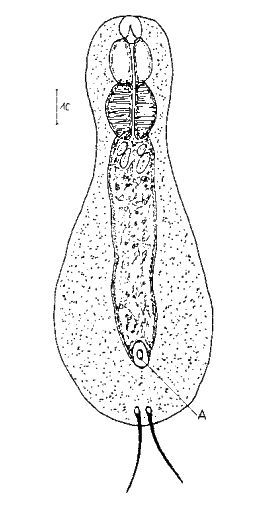

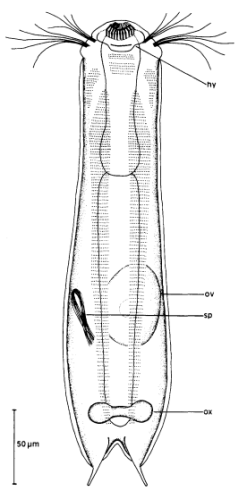
 typische Schuppen eines Aspidiophorus, hier A. squamulosus
typische Schuppen eines Aspidiophorus, hier A. squamulosus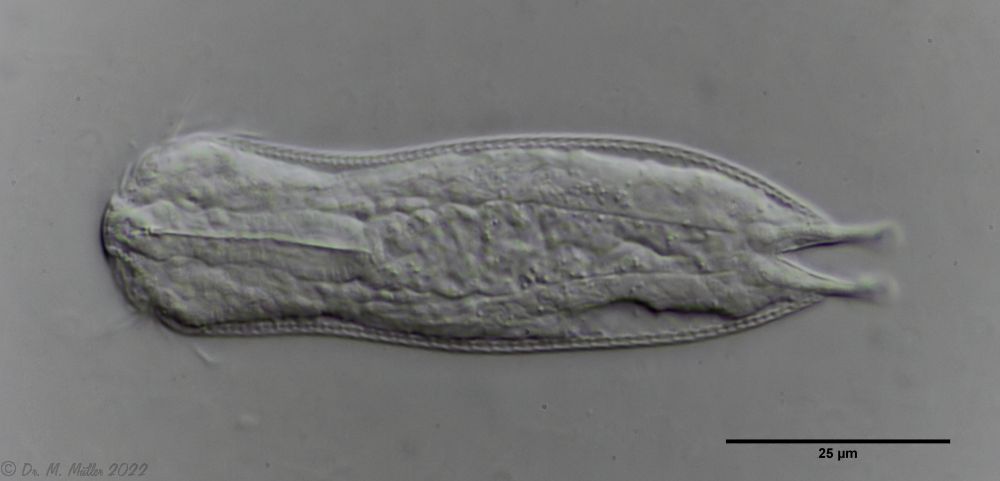
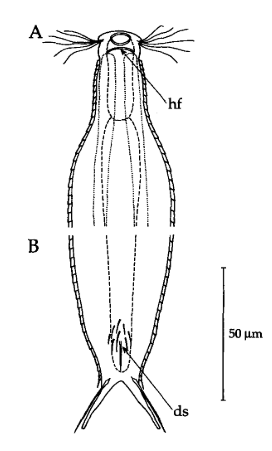
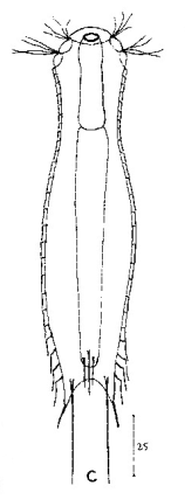
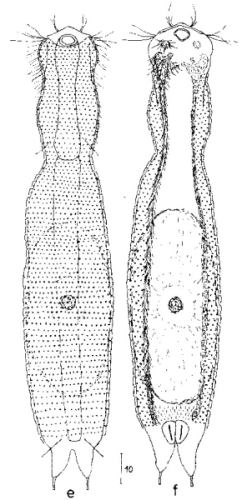
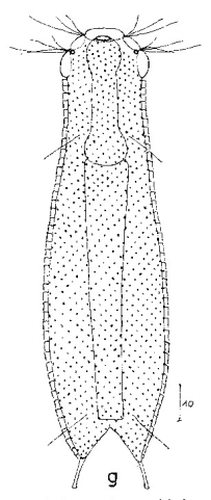
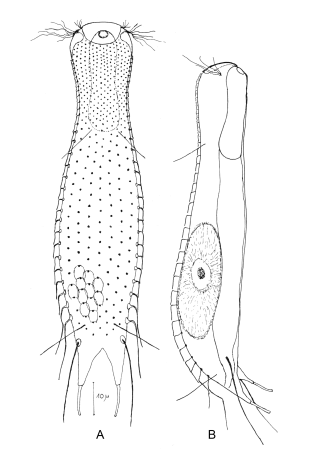
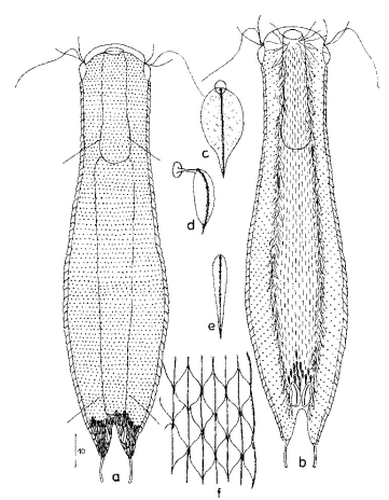
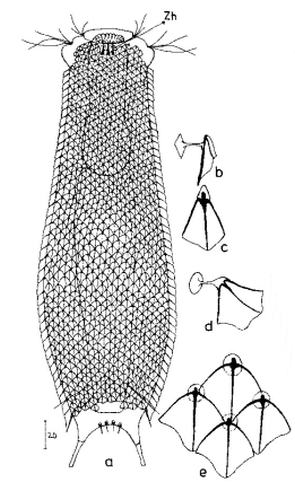
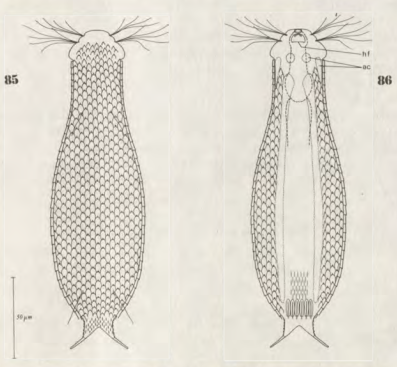
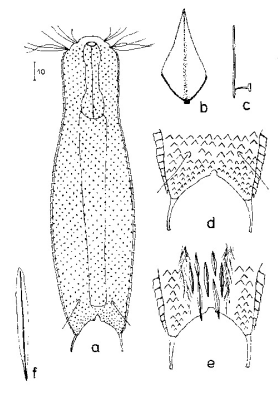
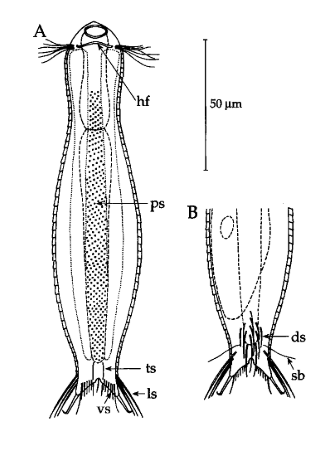
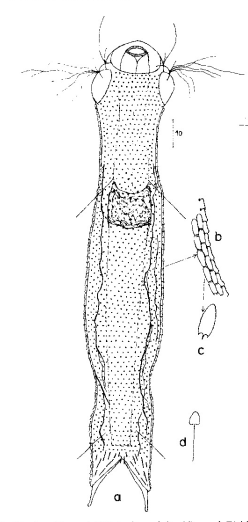
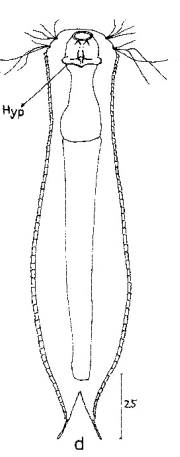
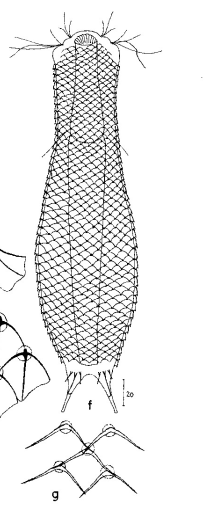
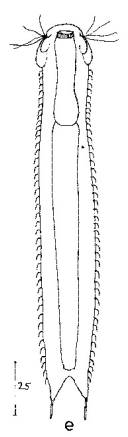
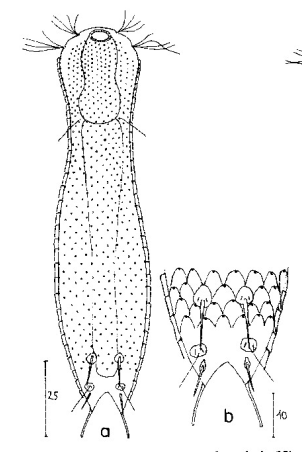
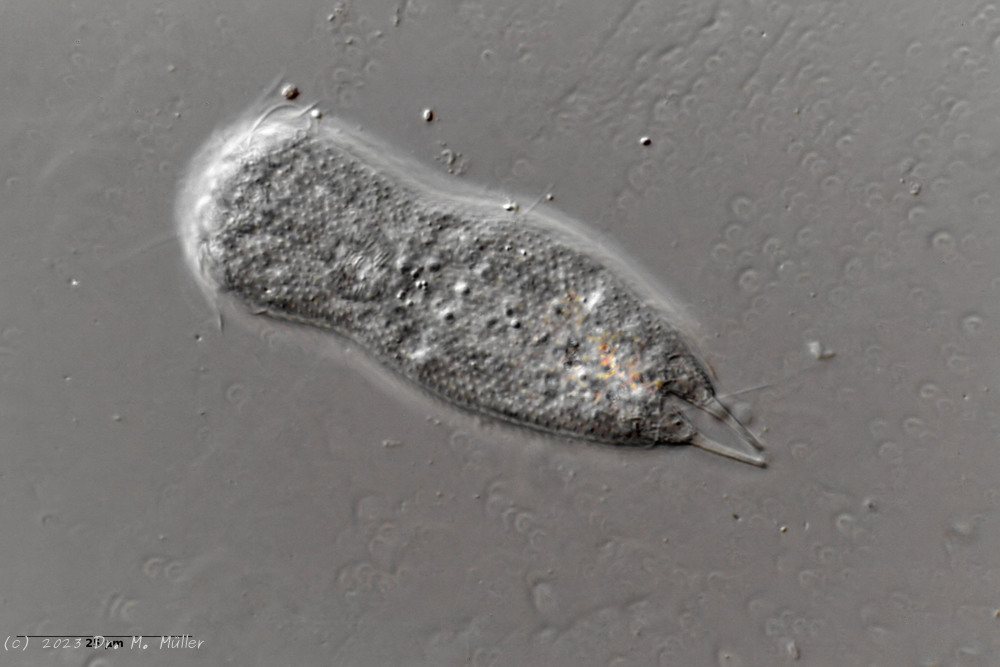
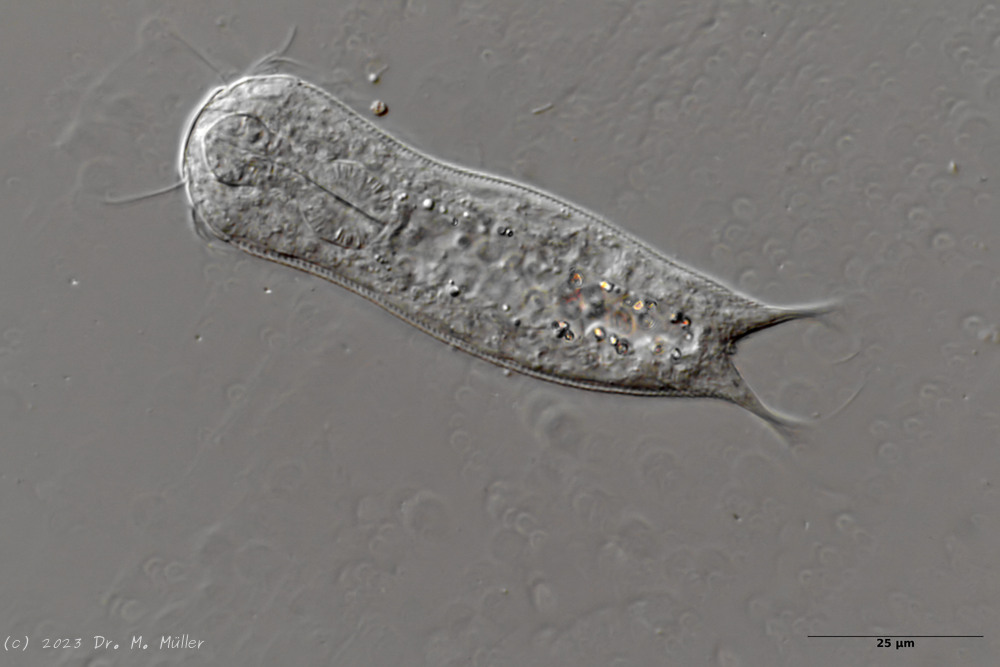
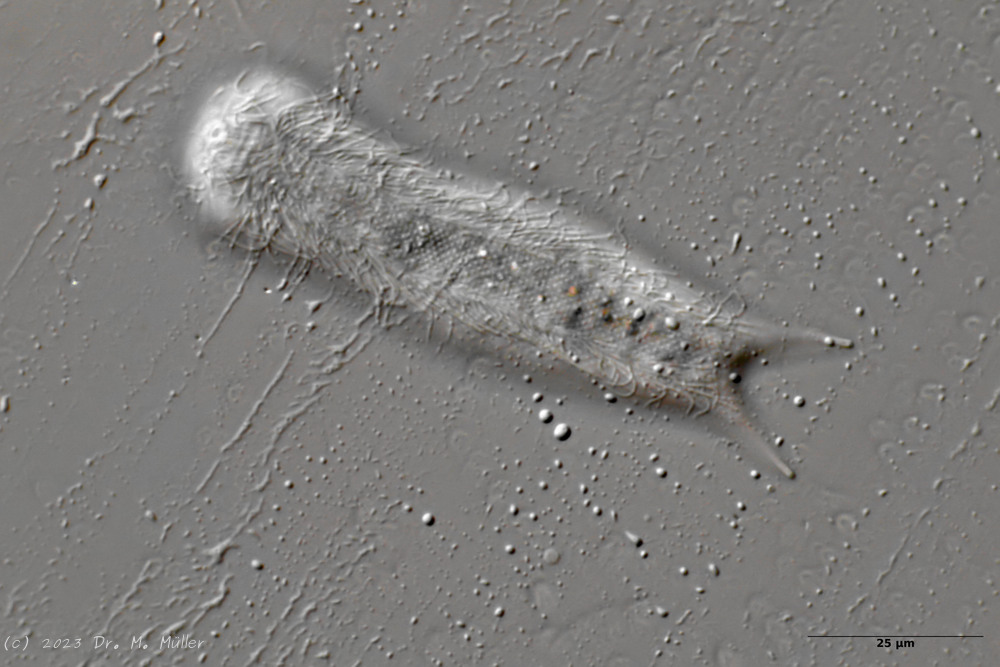
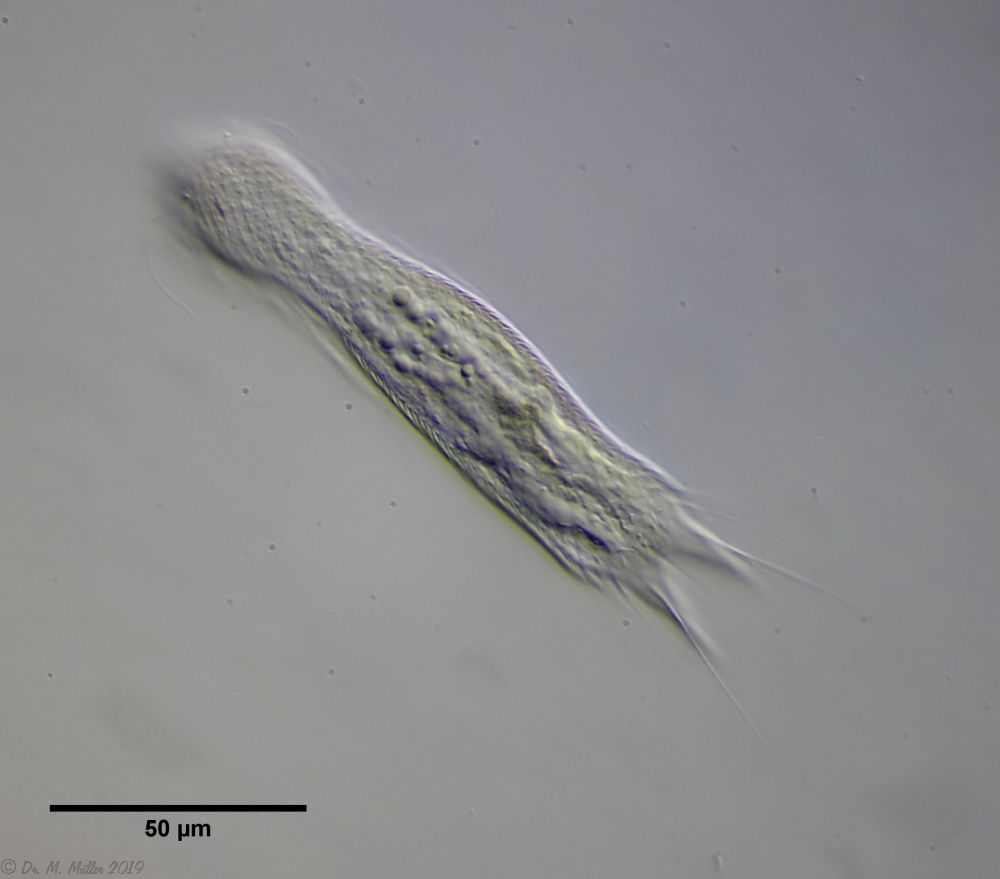


 Rückenschuppen
Rückenschuppen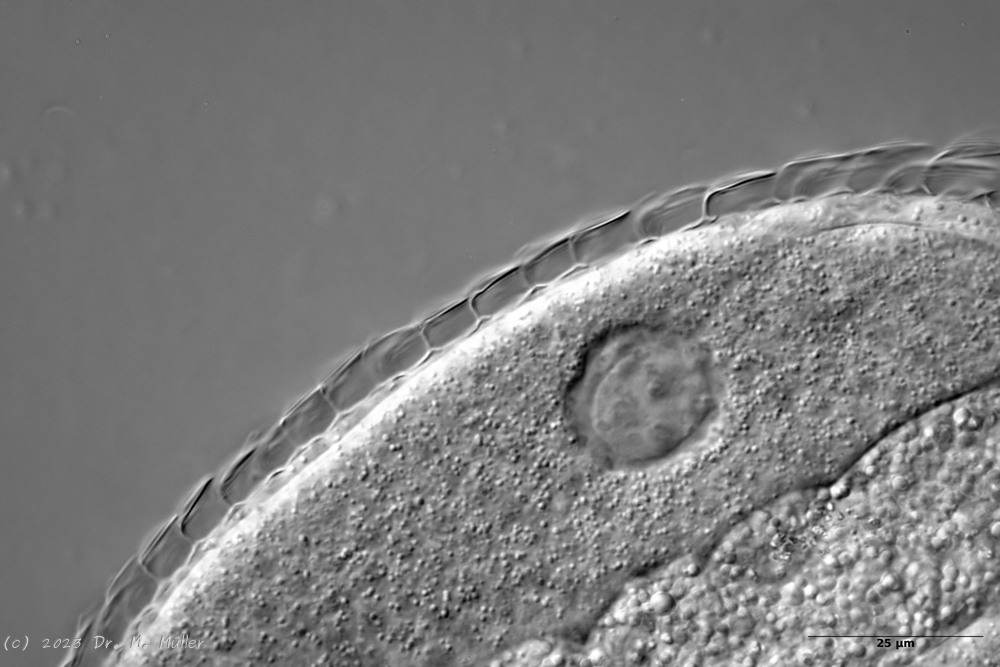
 Querschnitt Schuppenstiele
Querschnitt Schuppenstiele

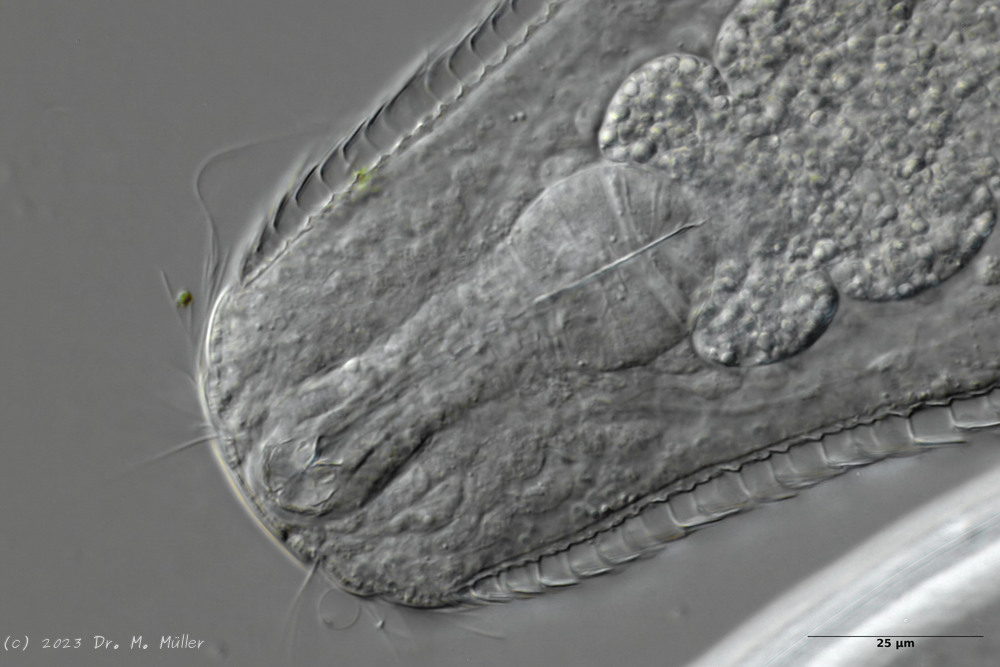
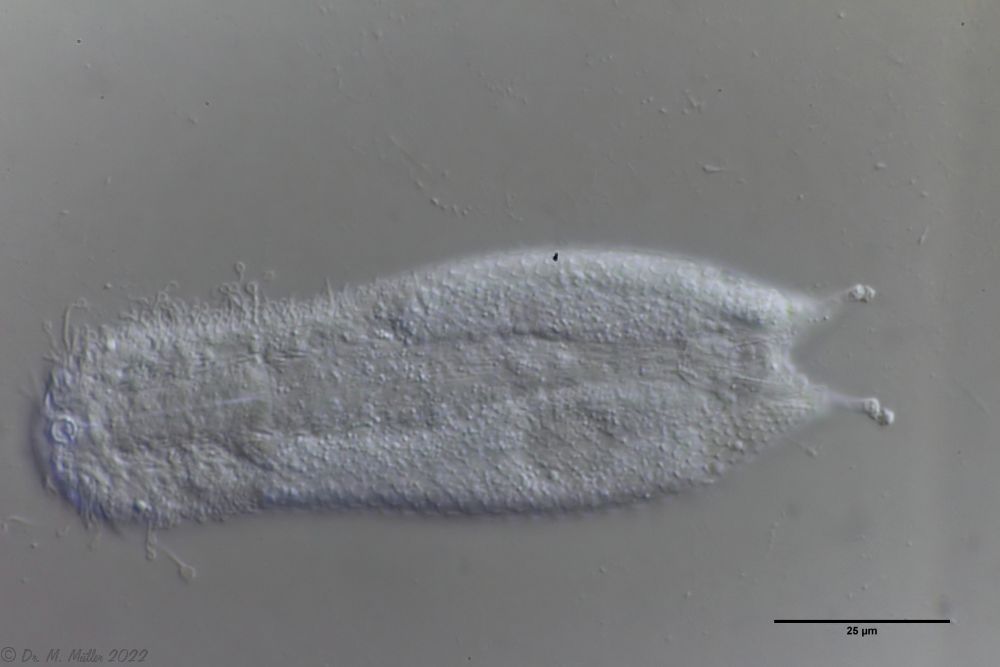
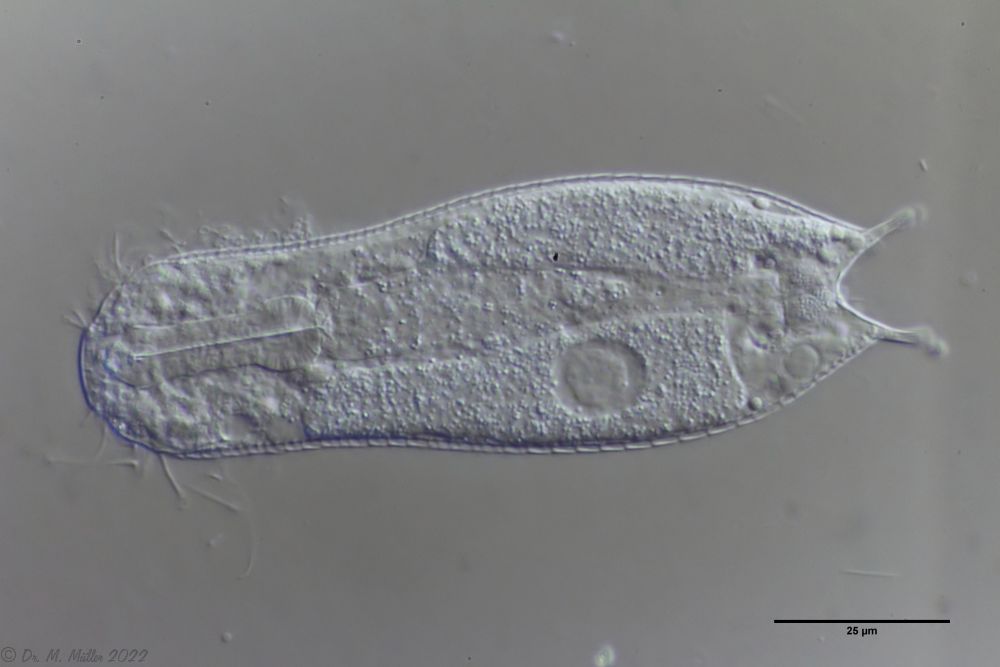
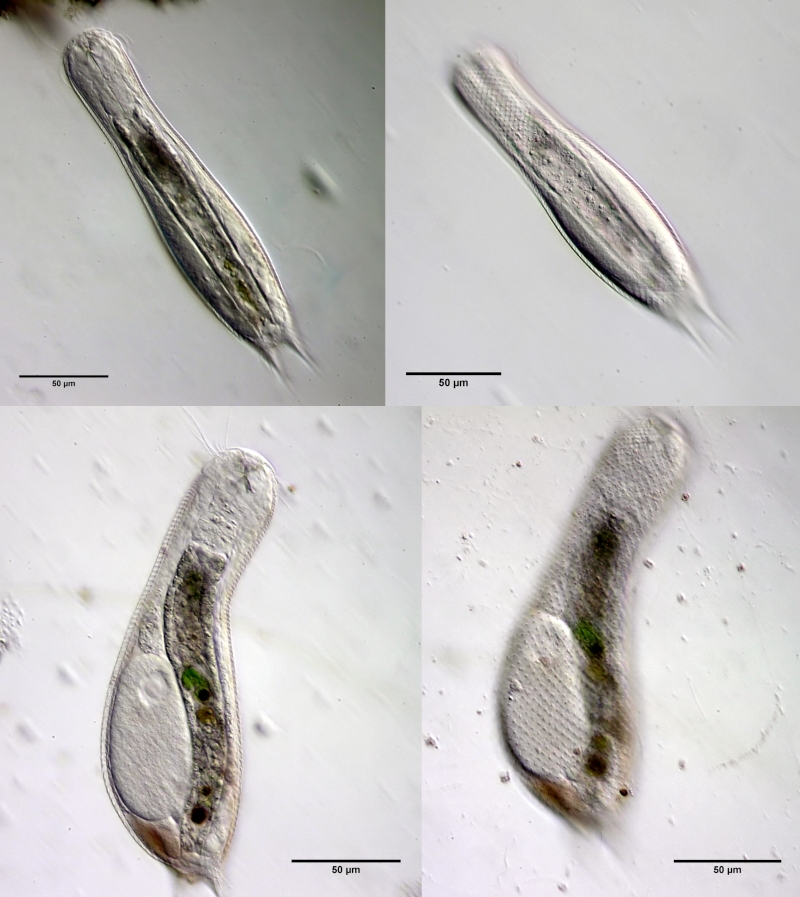
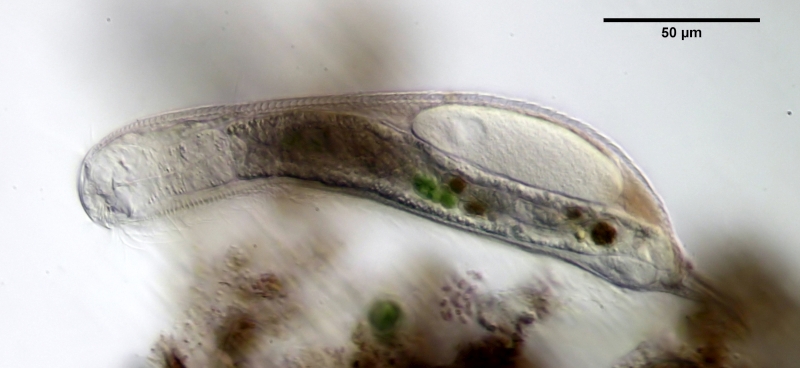

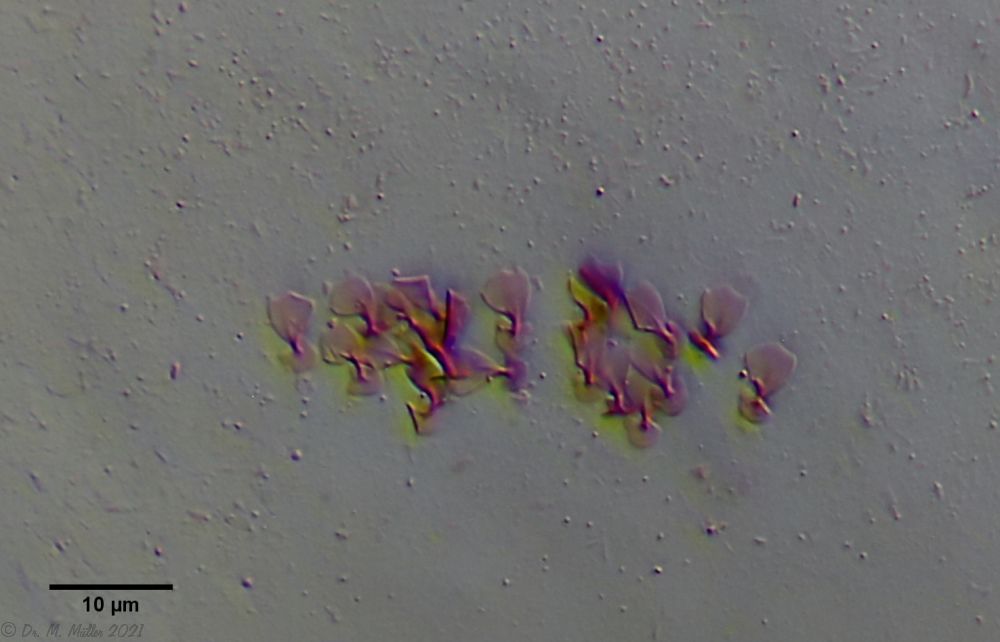
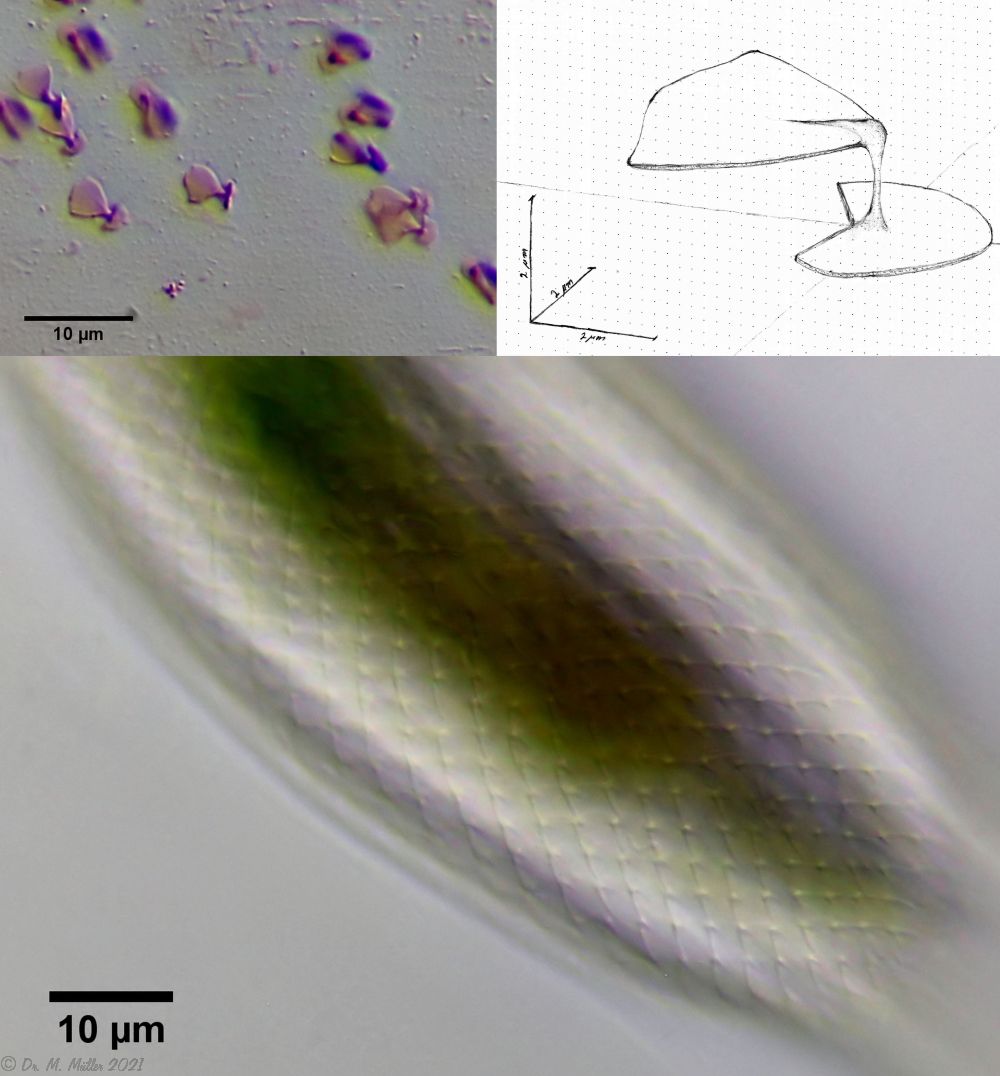
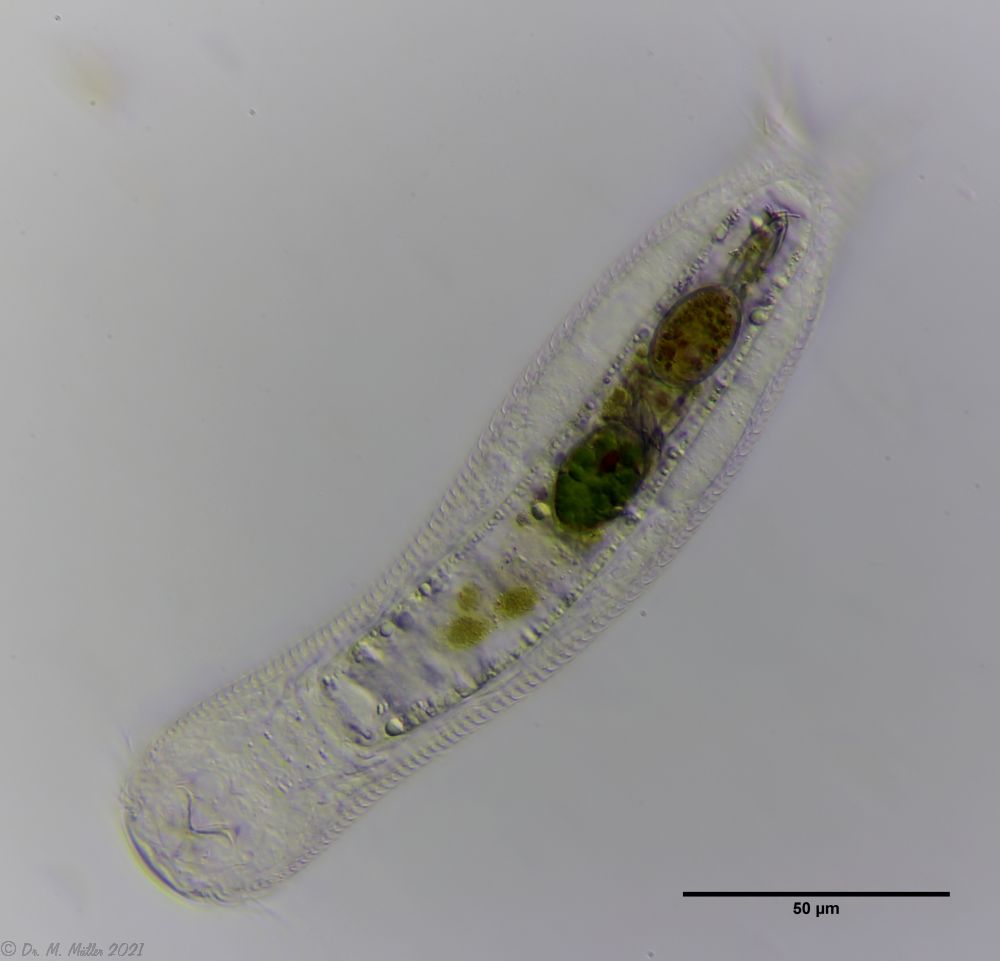
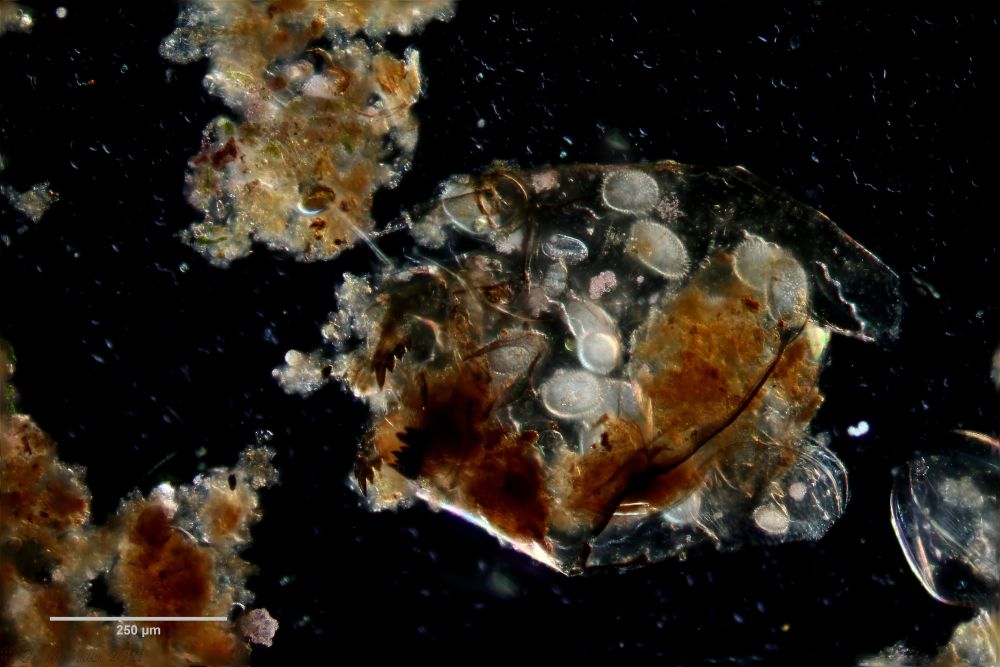
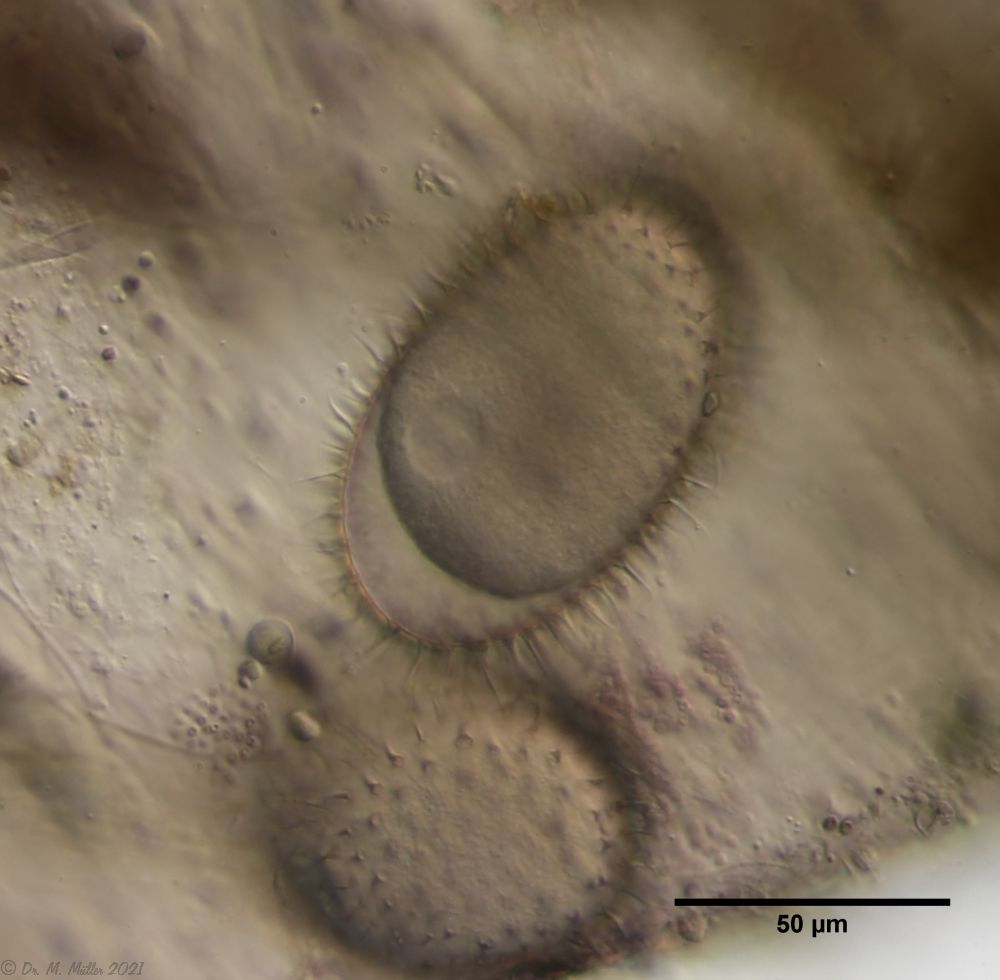

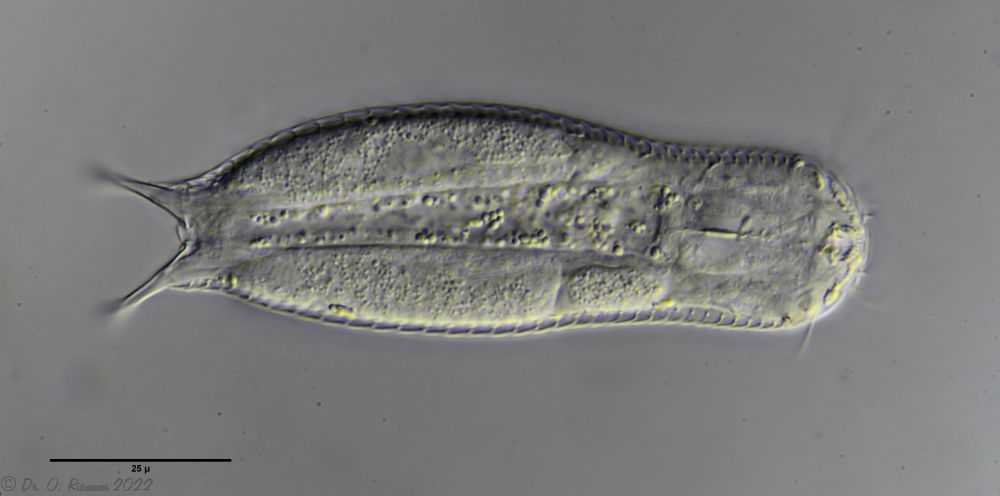
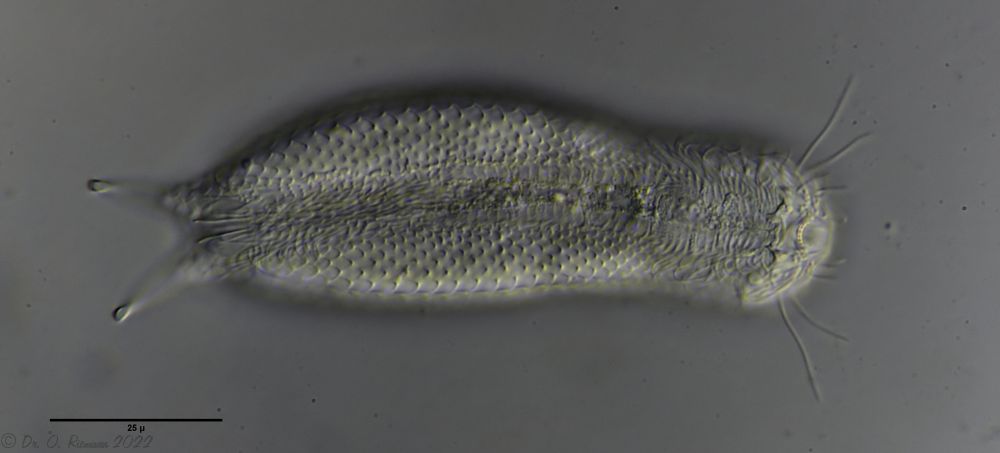
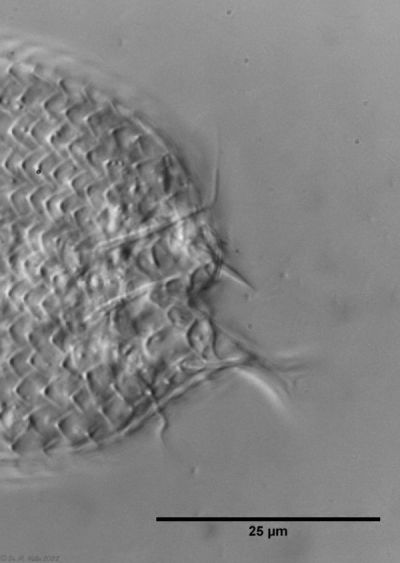
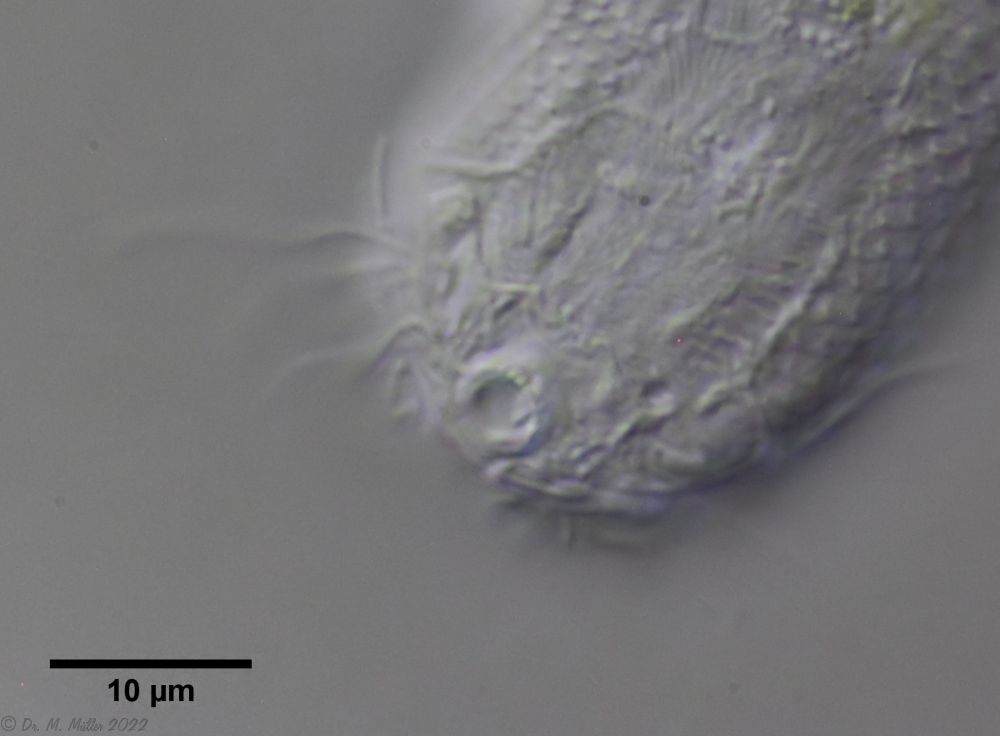
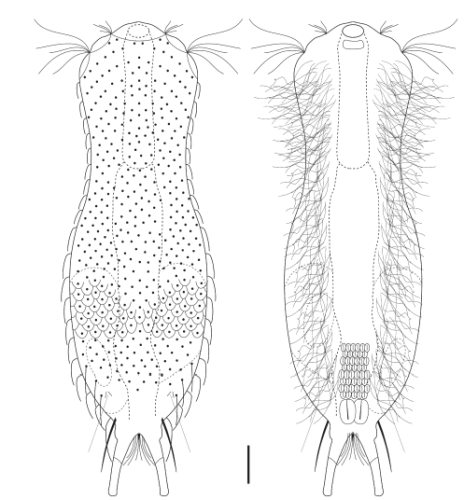
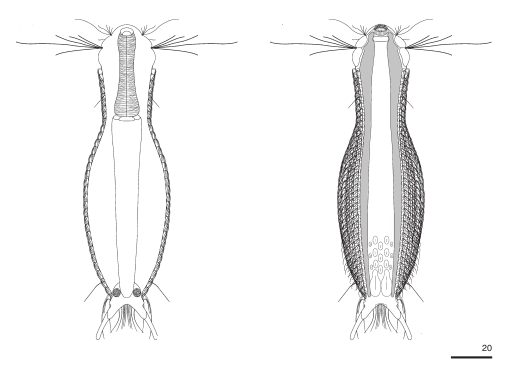
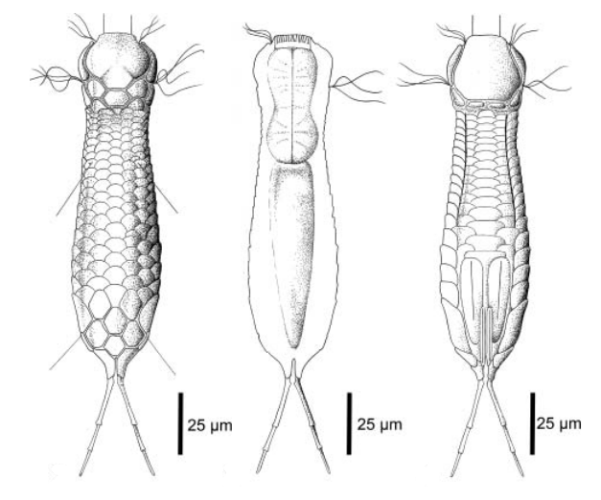
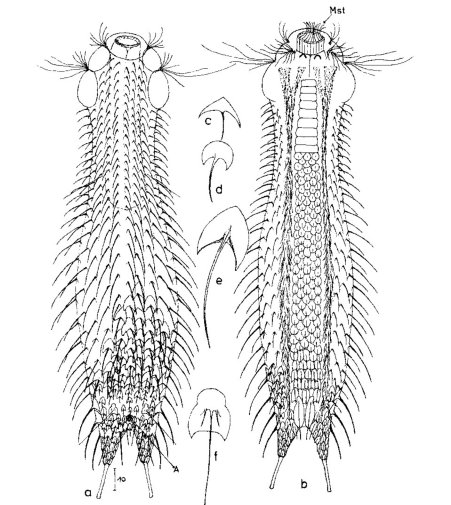
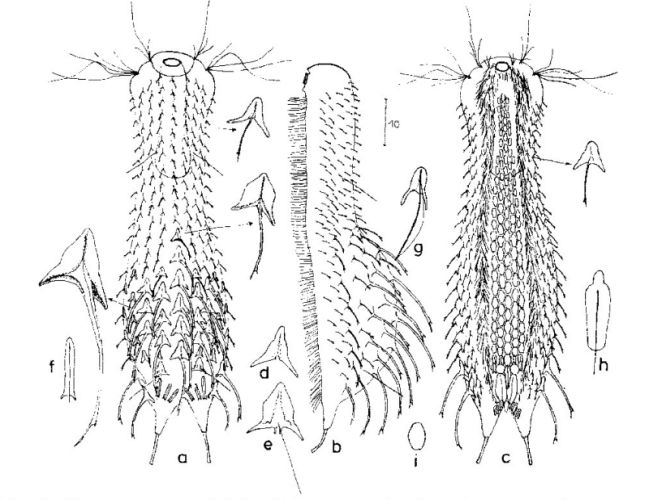
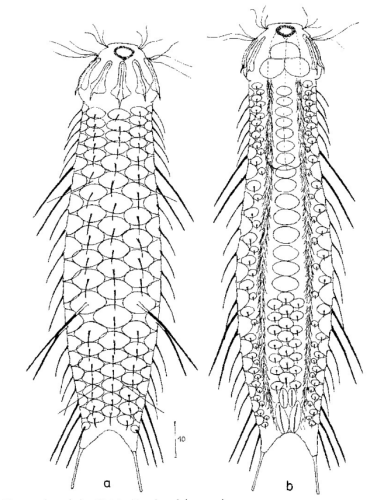
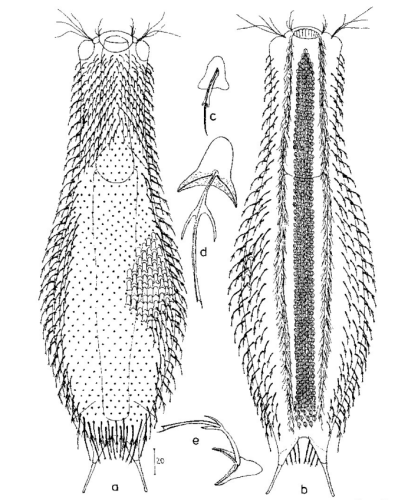
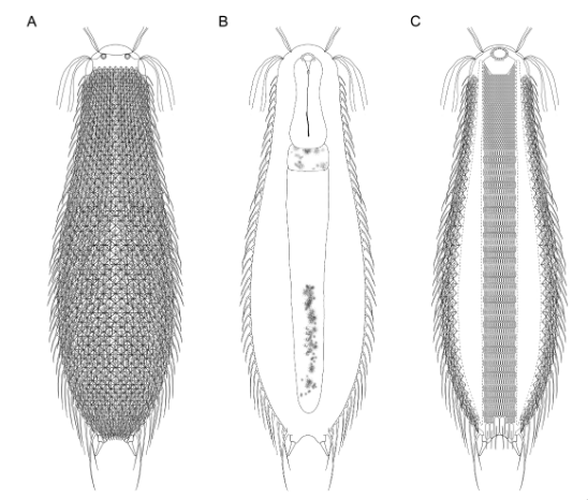
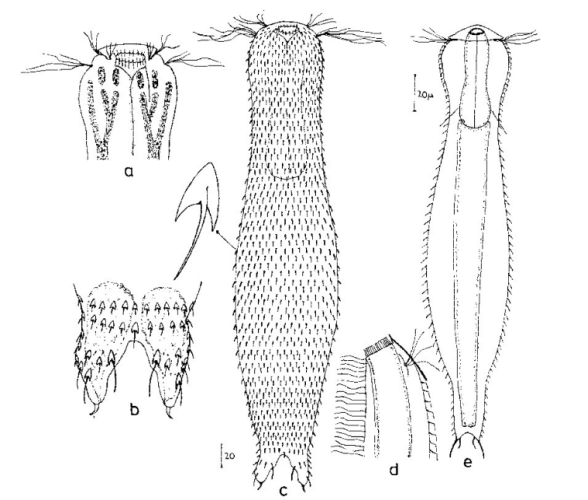
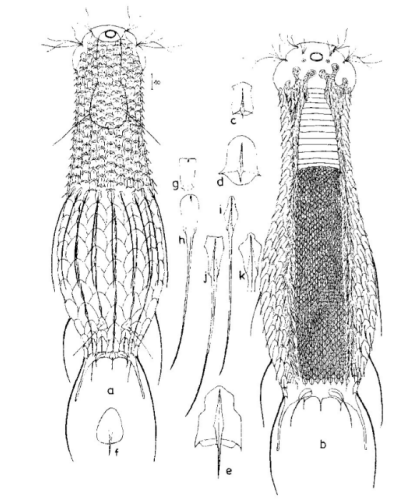
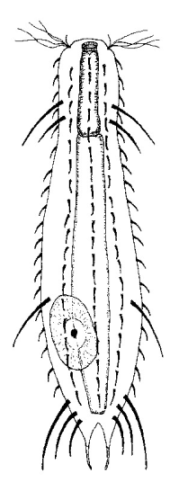
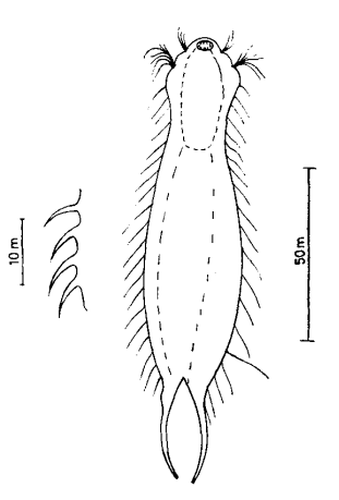
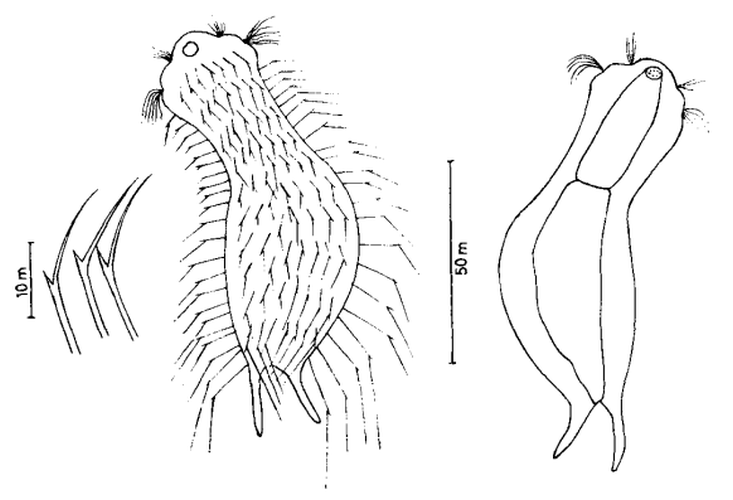
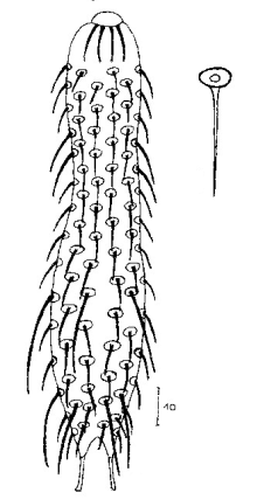
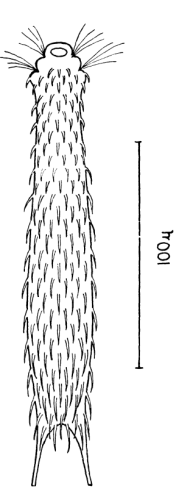
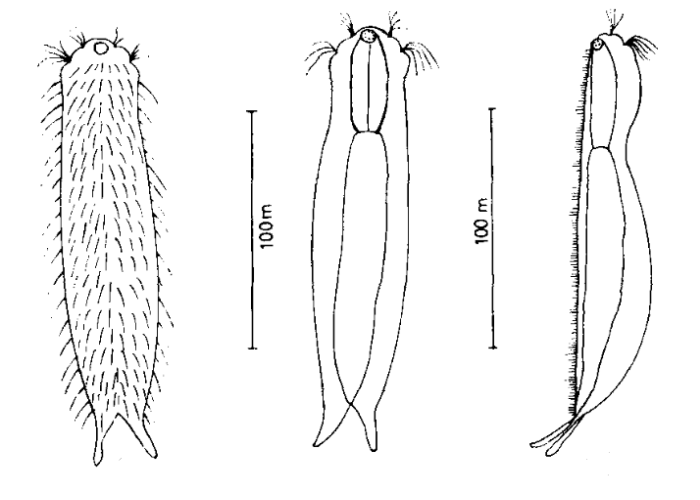
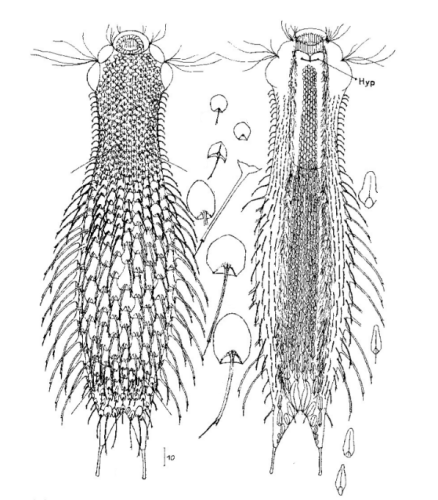
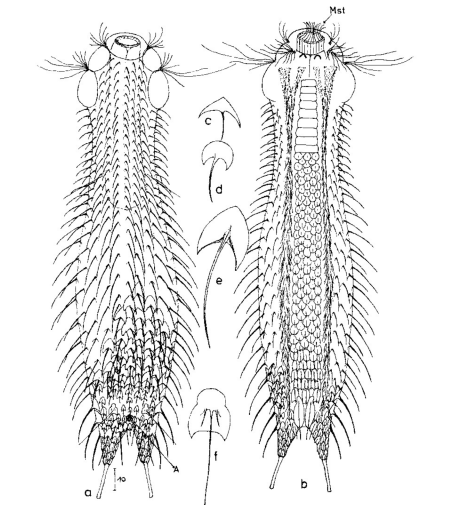
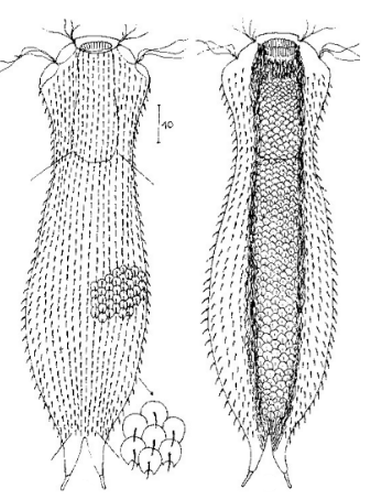
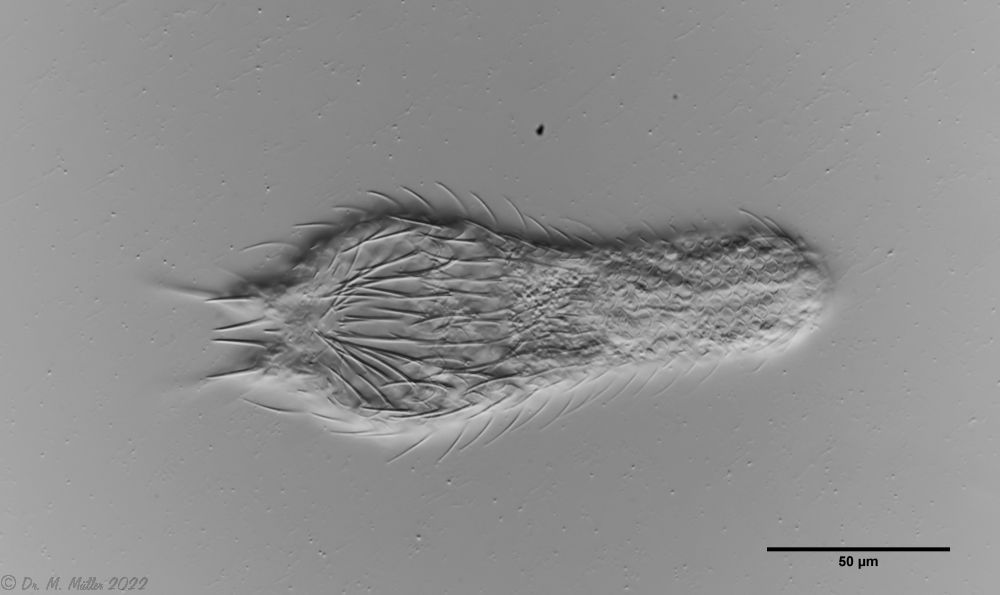
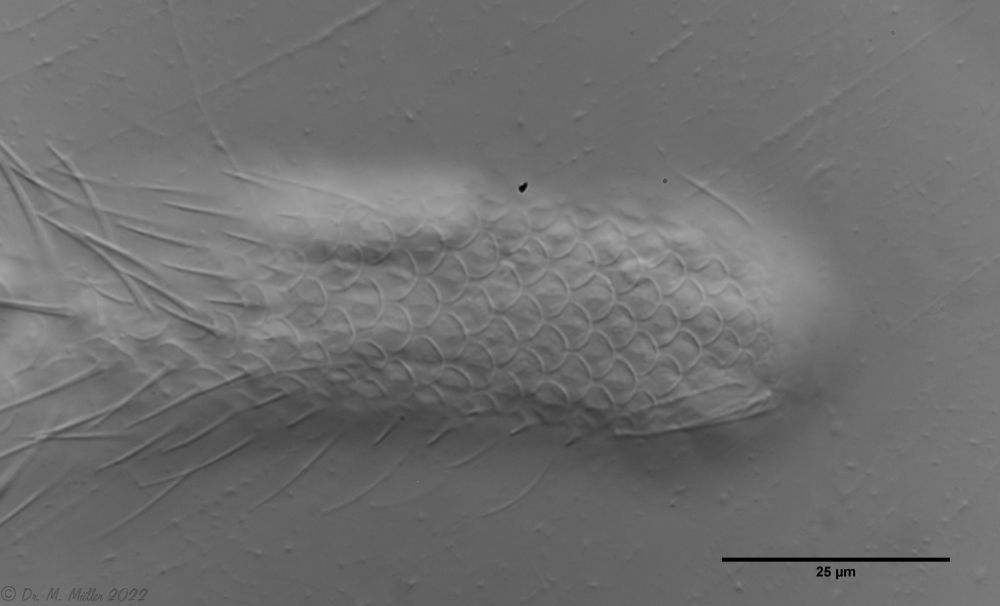
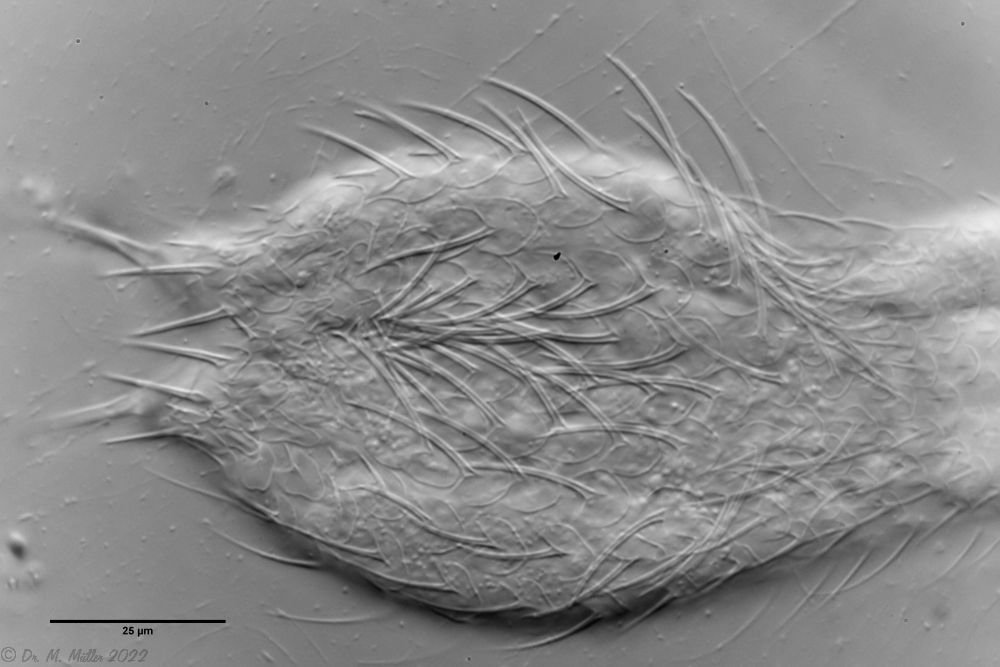

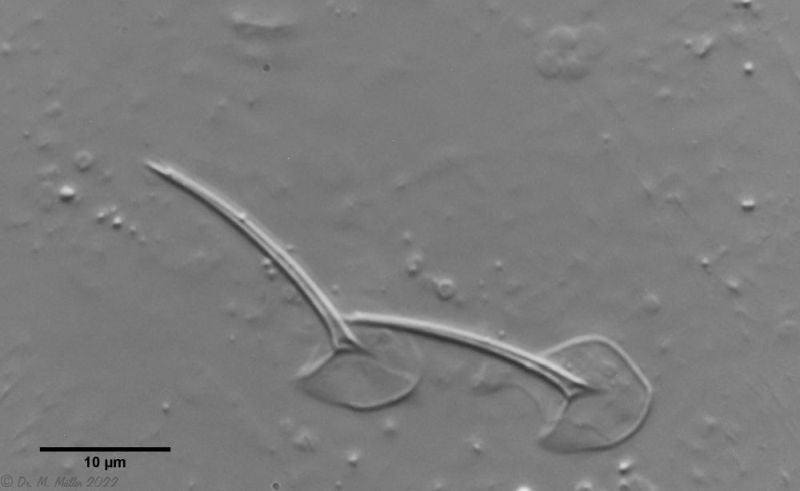
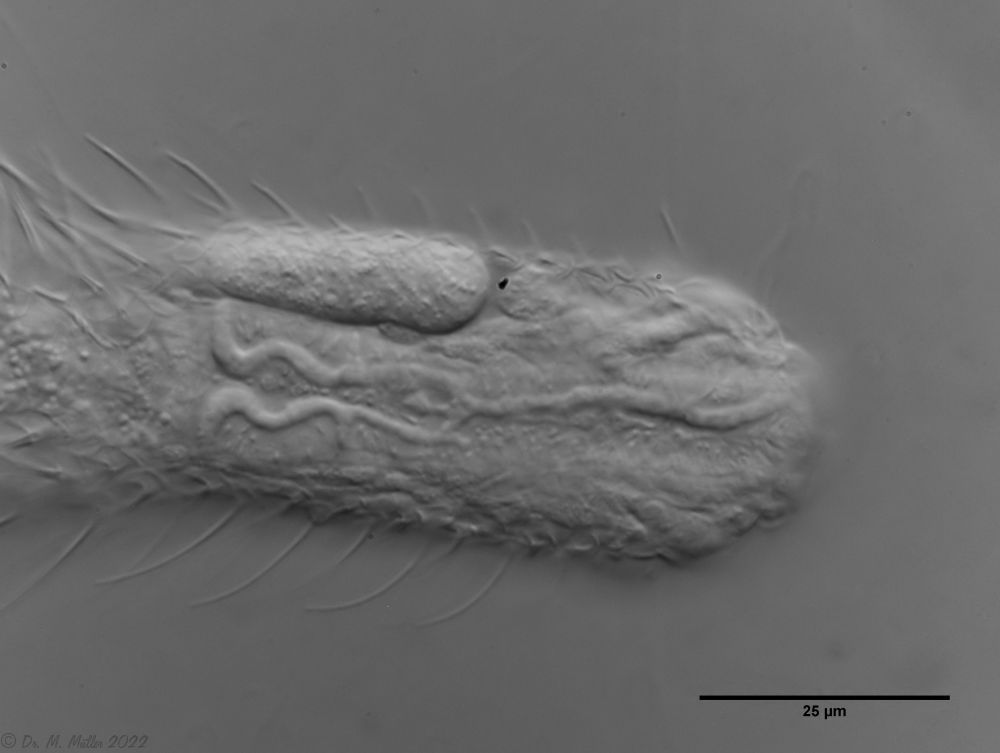
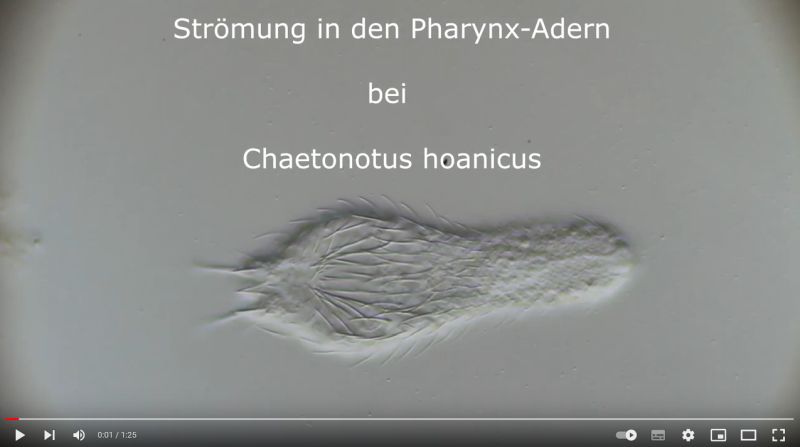
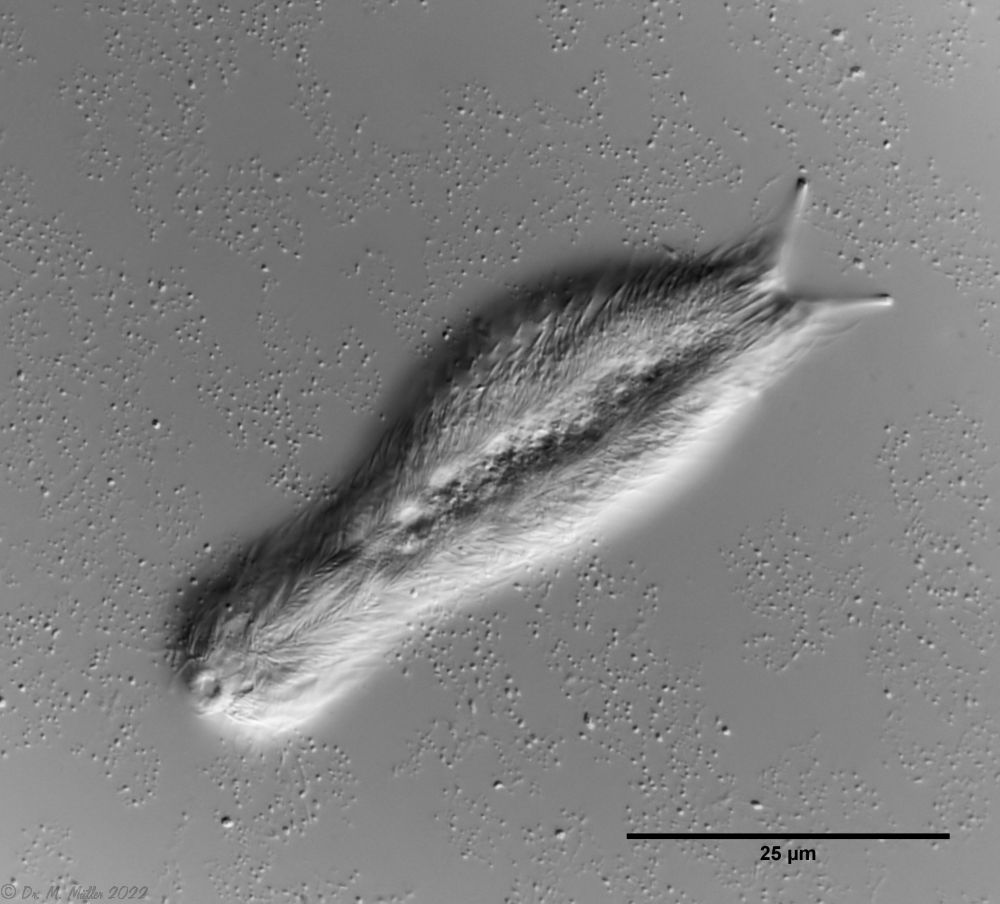
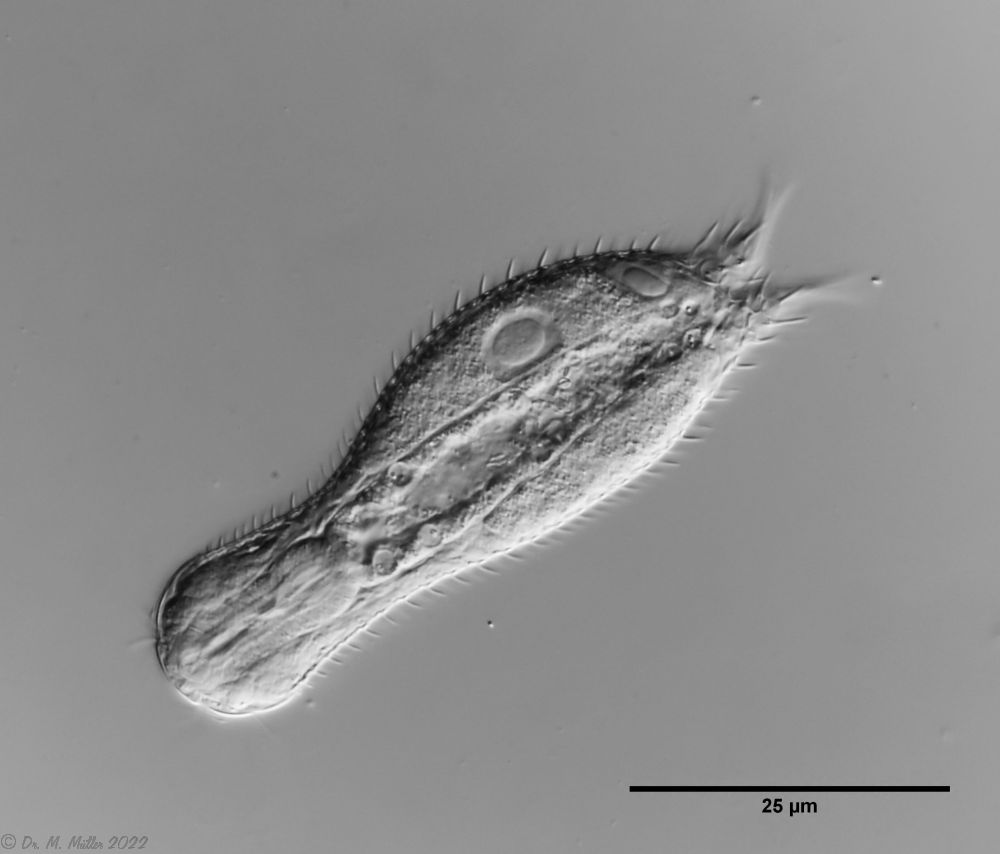
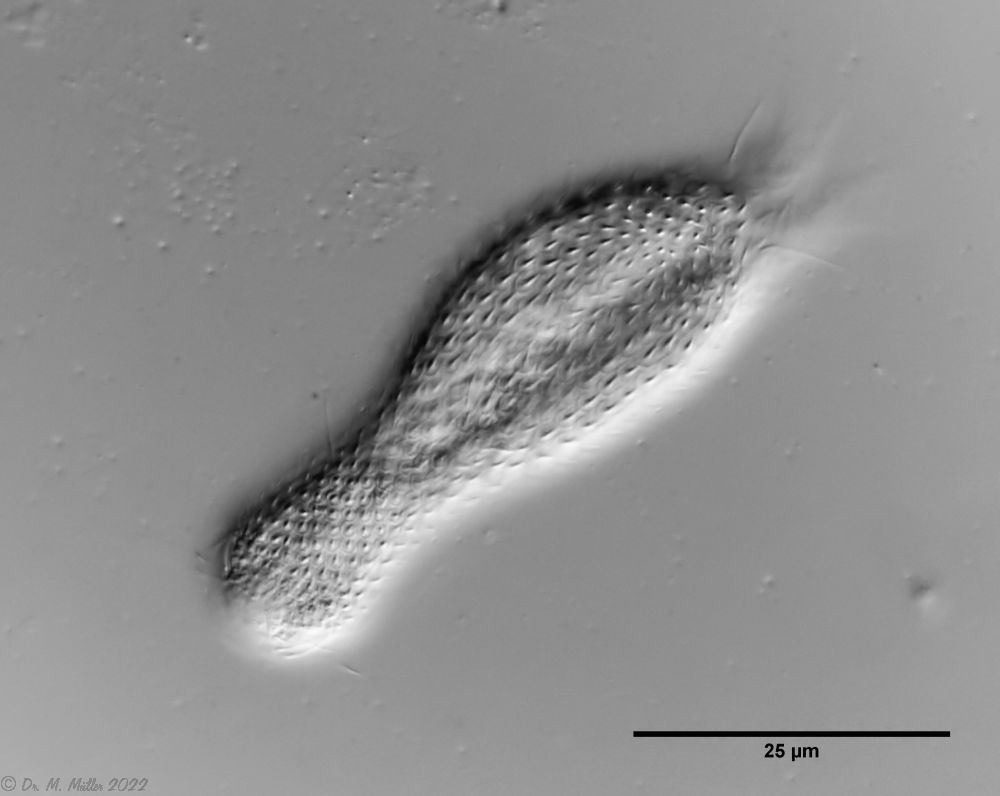

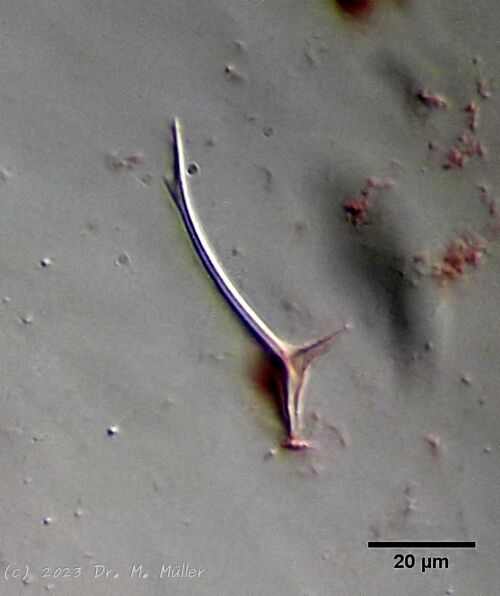
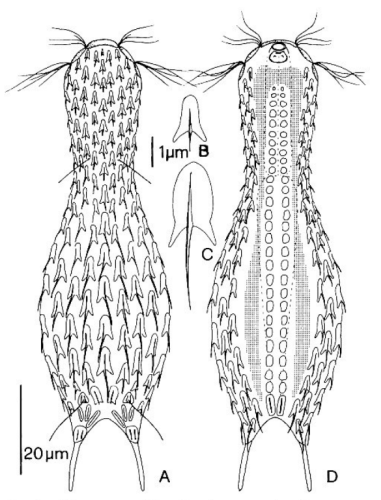
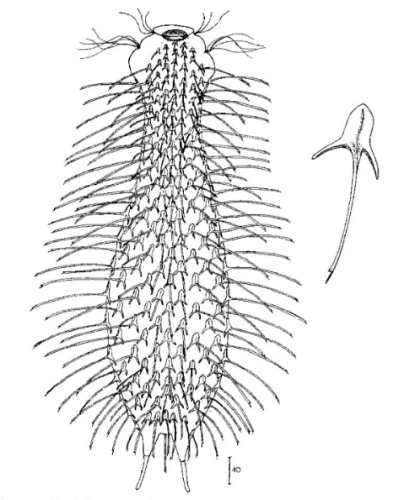
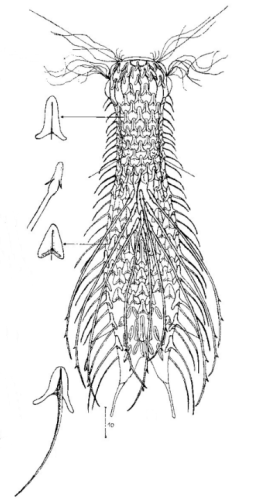
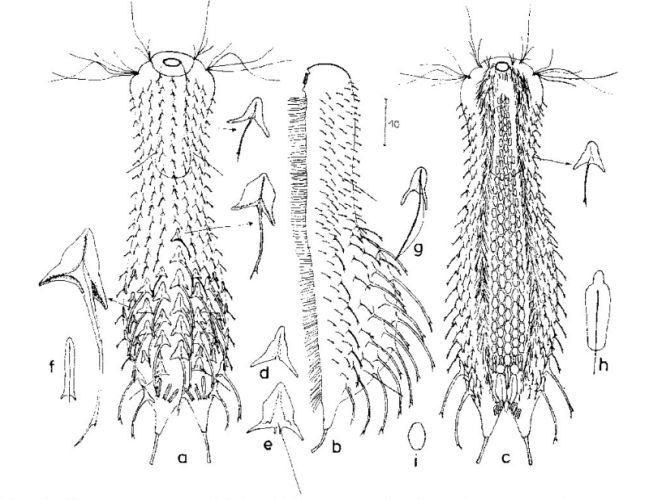
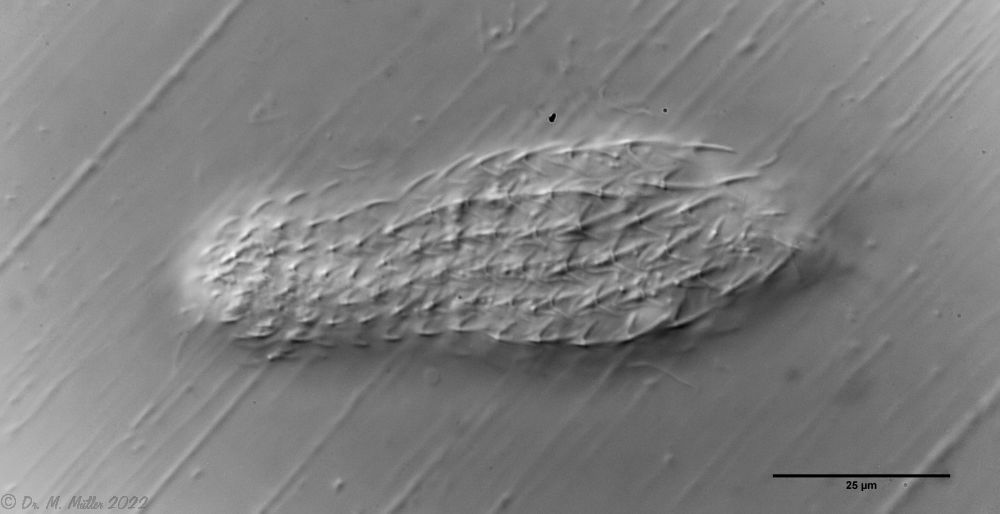
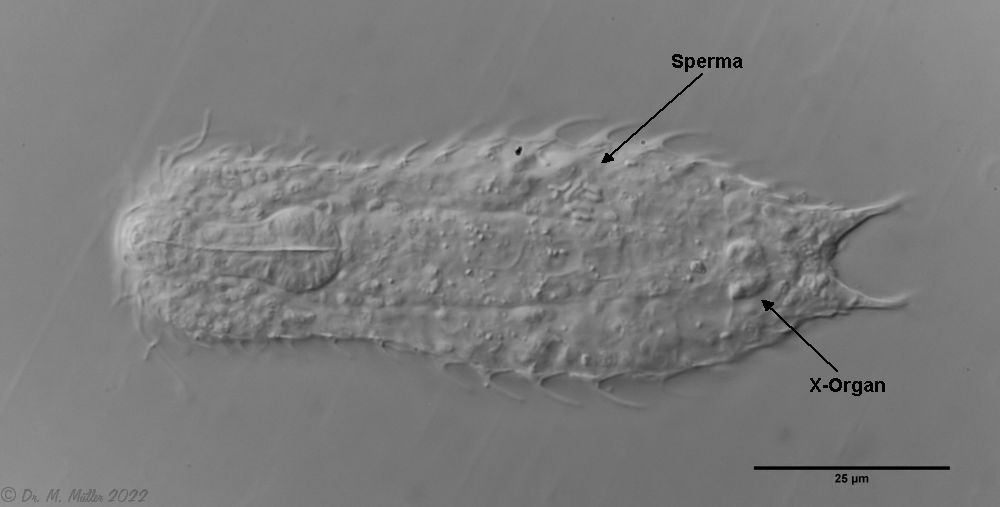
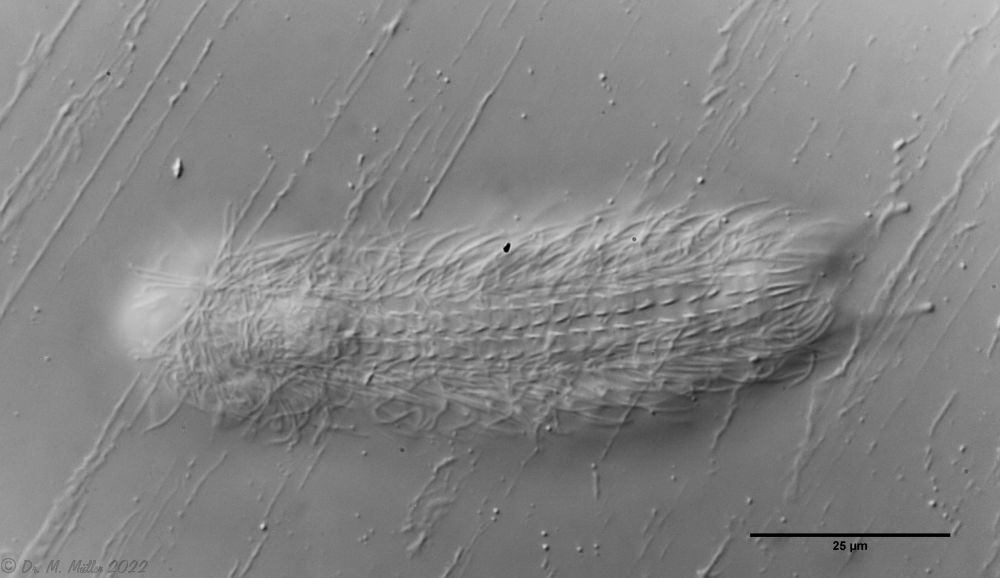
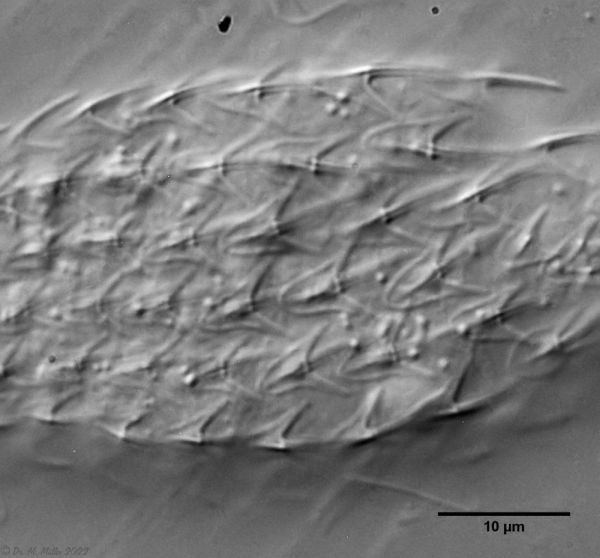
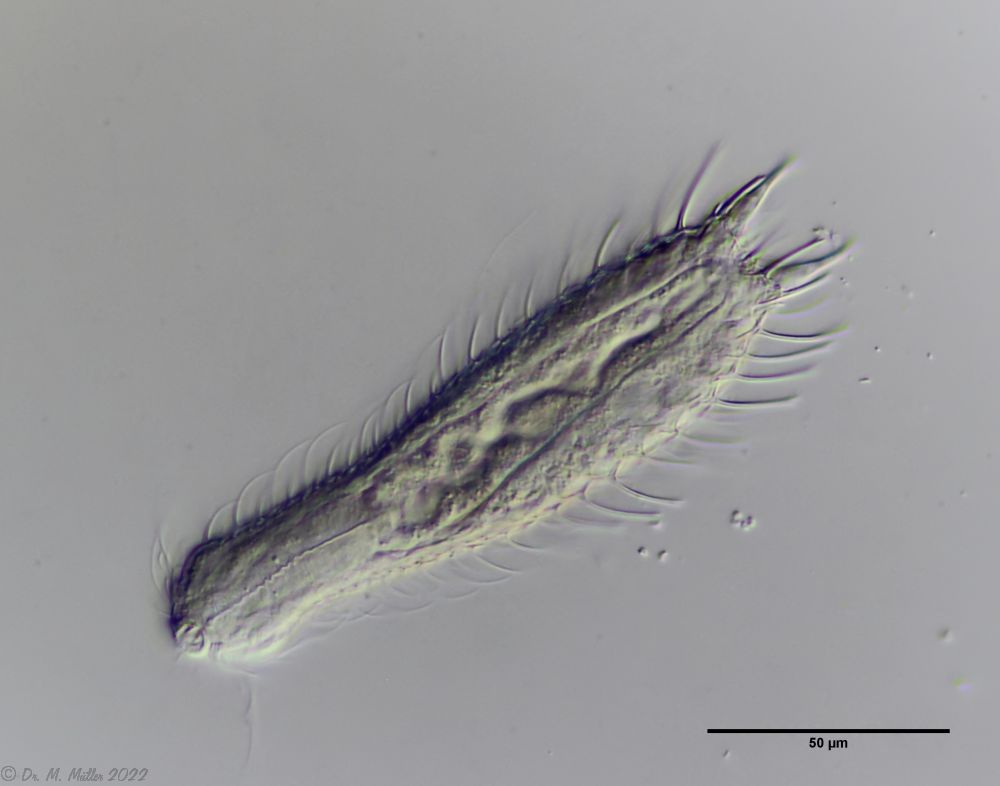



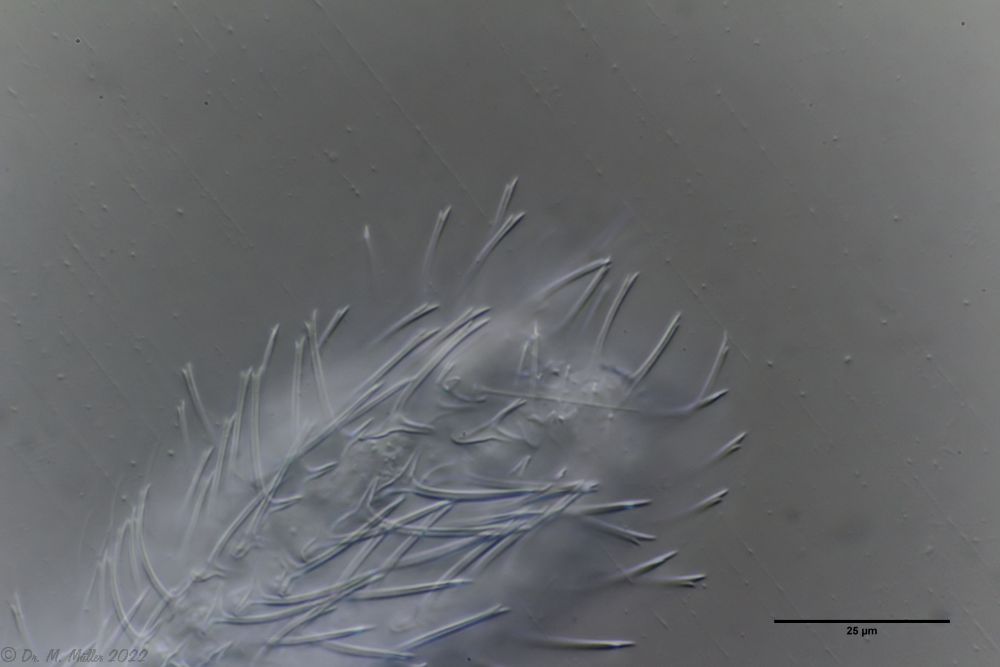 Ch. murrayi: dorsale Beschuppung des Hinterendes
Ch. murrayi: dorsale Beschuppung des Hinterendes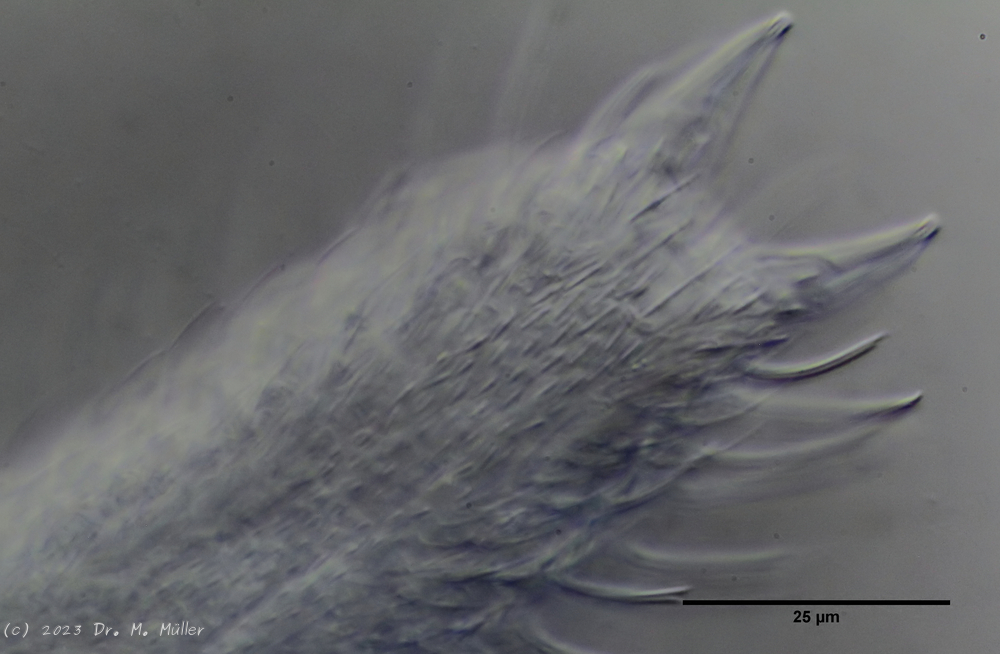 Ch. murray: Schuppen des ventrale Zwischenfeldes
Ch. murray: Schuppen des ventrale Zwischenfeldes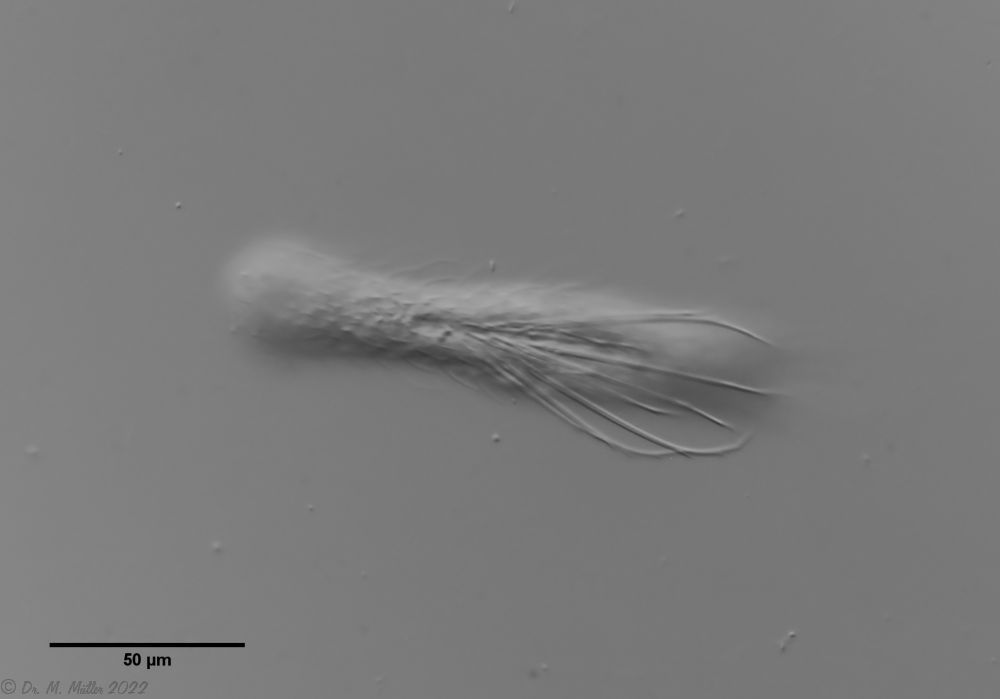
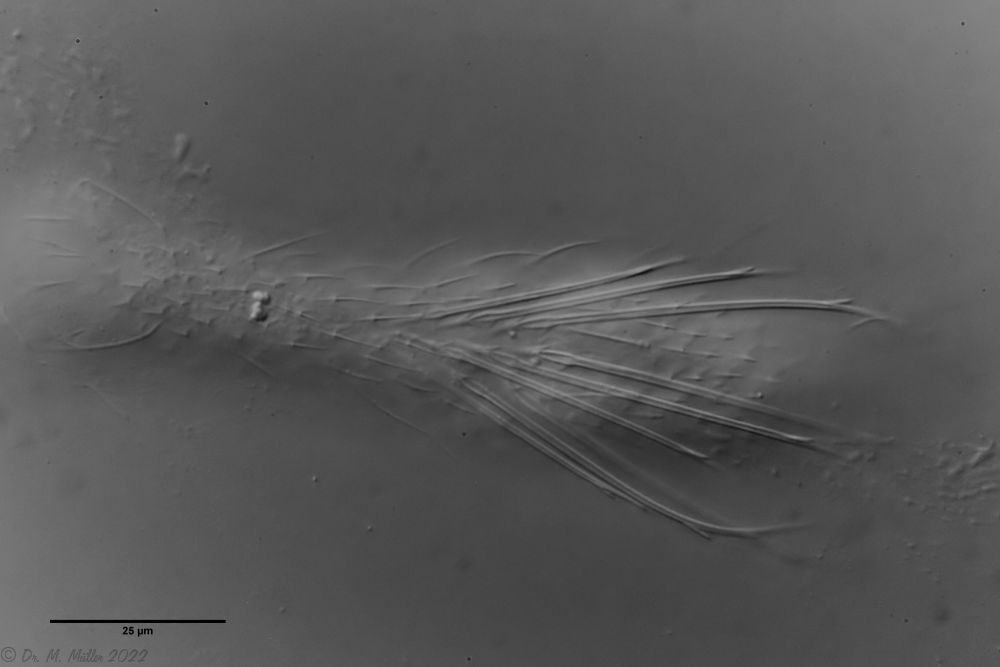
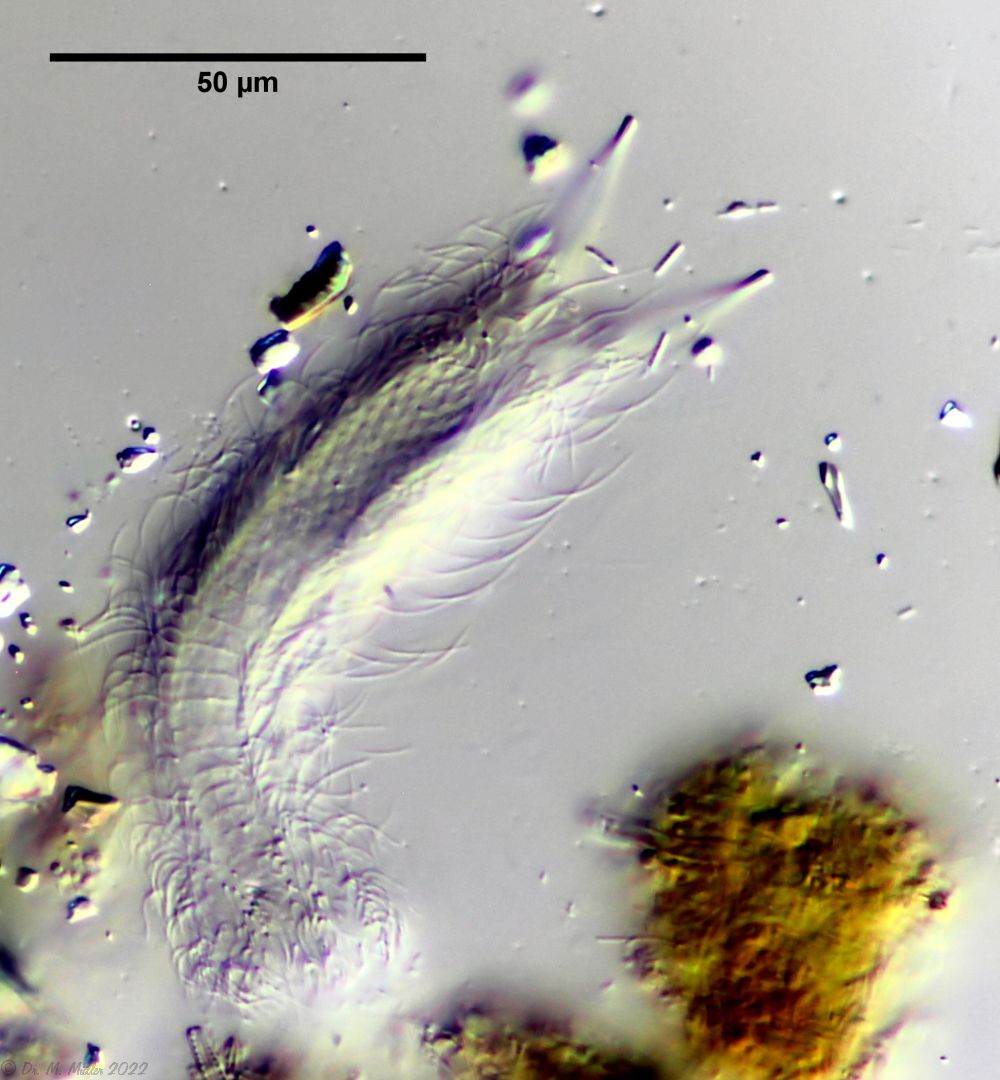

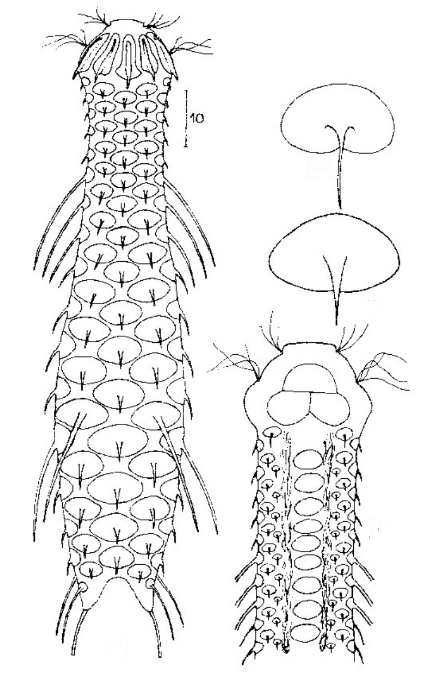

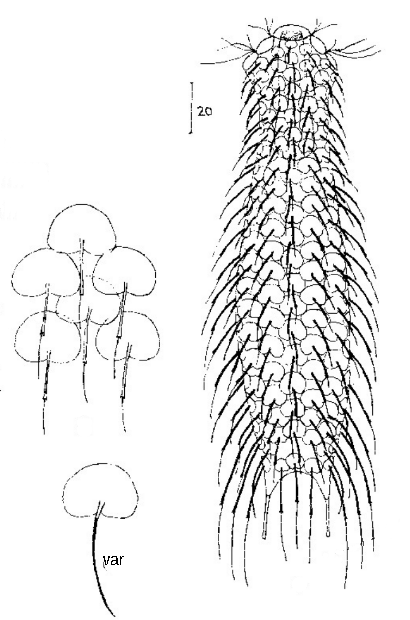

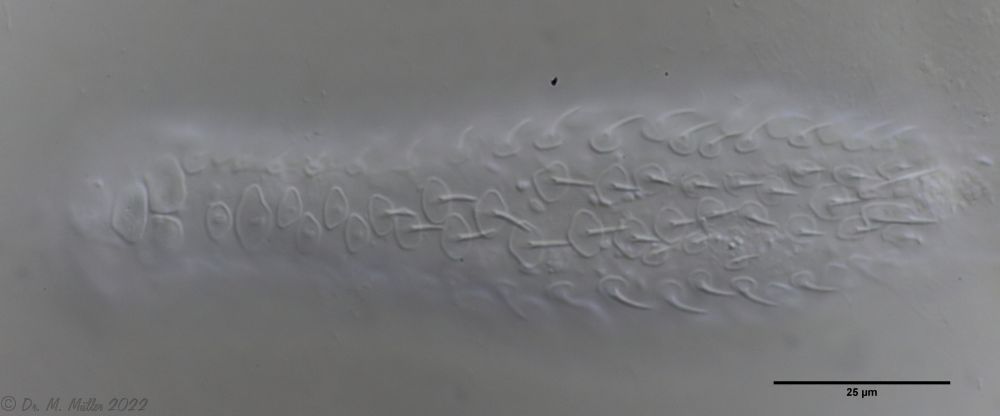 Ch. acanthocephalus: ventral
Ch. acanthocephalus: ventral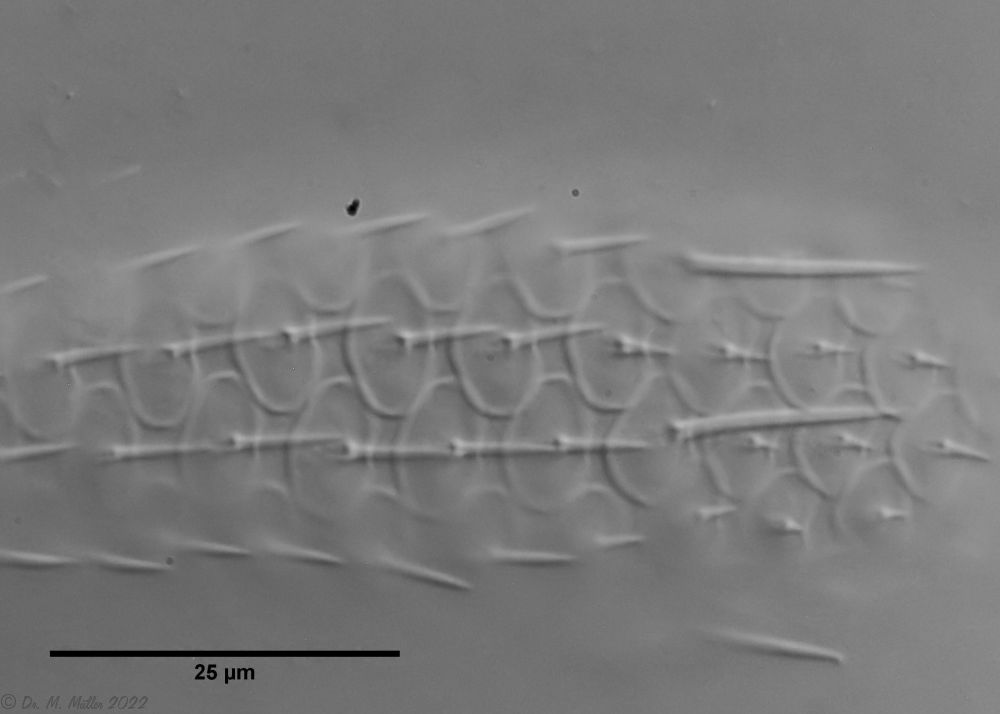

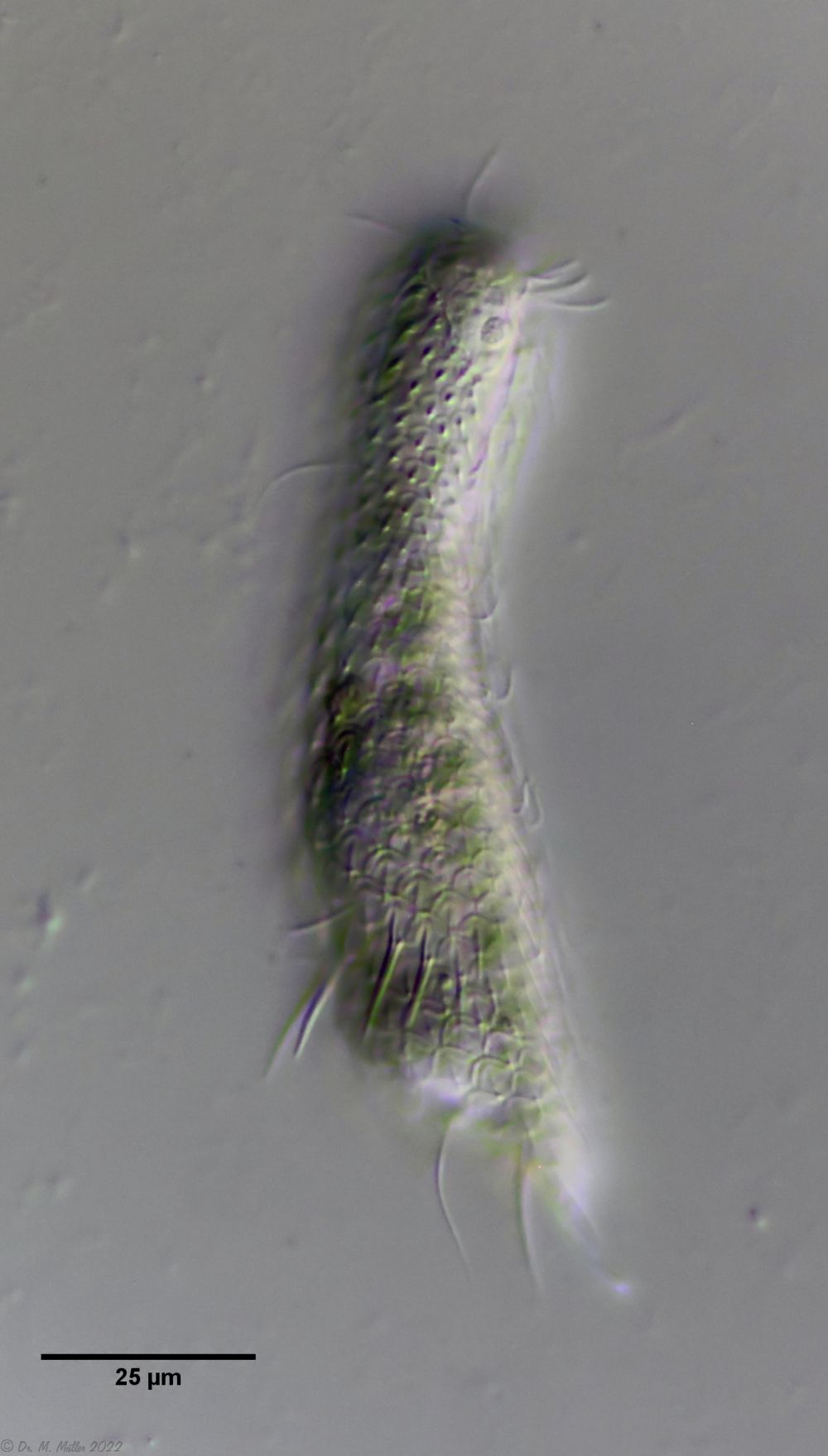
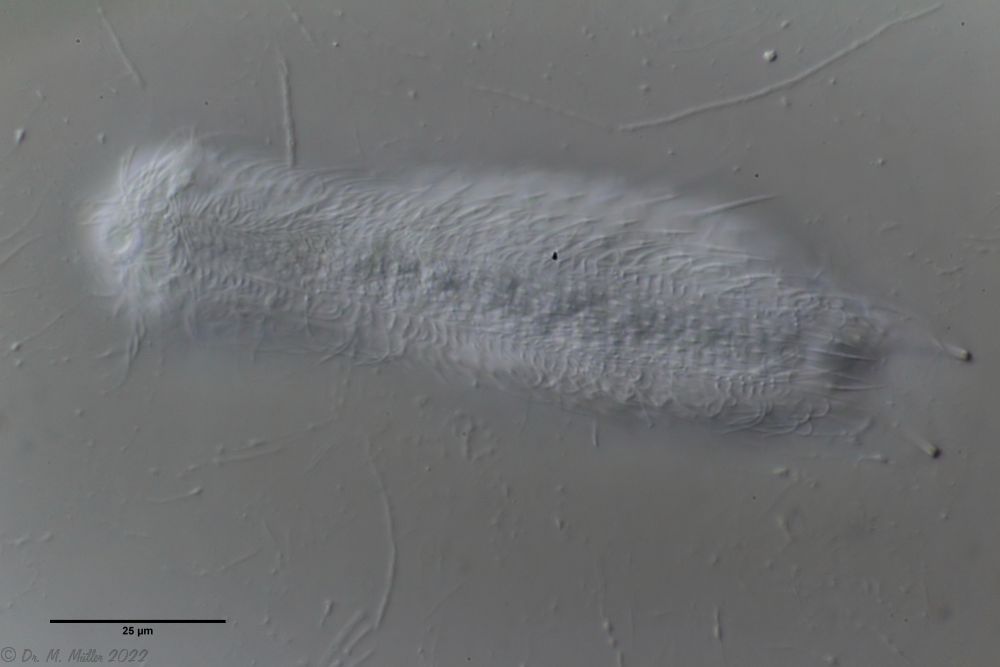
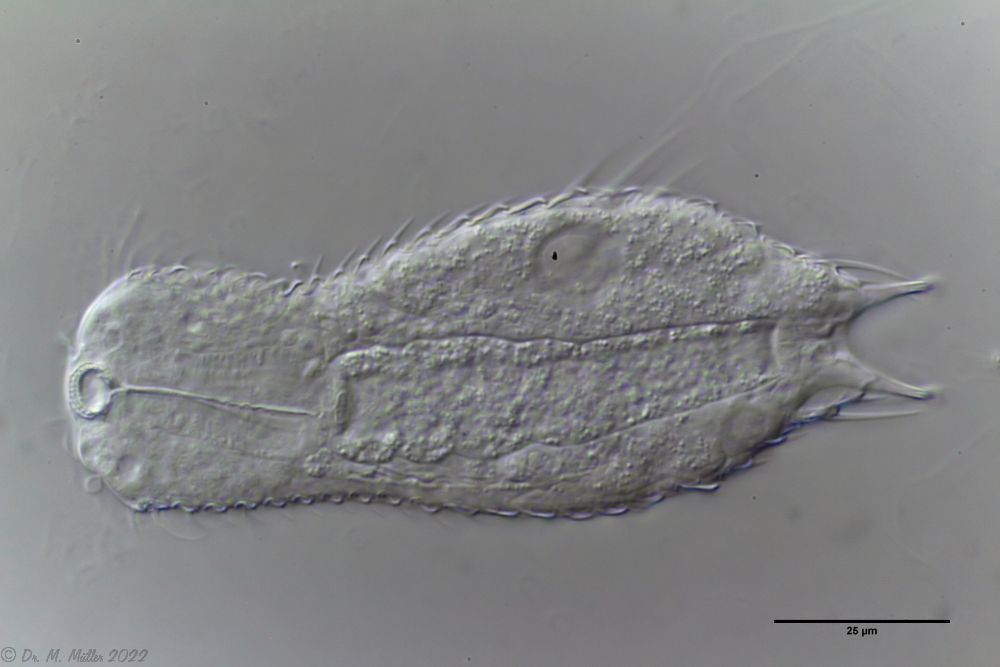
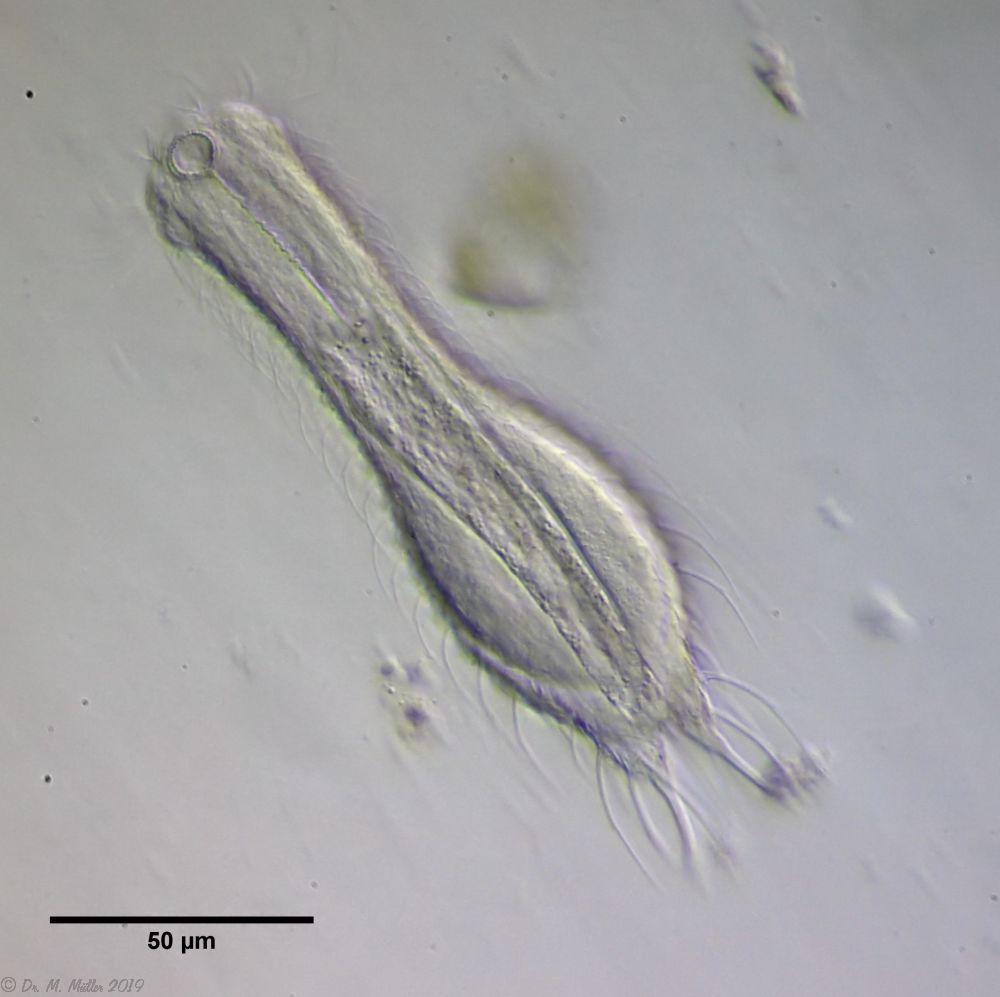
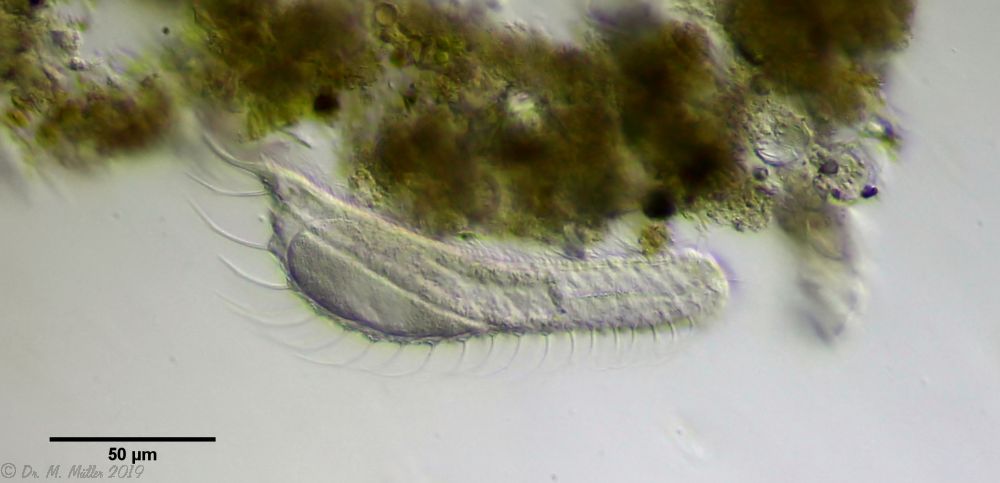 Ch. cordiformis: Seitenansicht
Ch. cordiformis: Seitenansicht
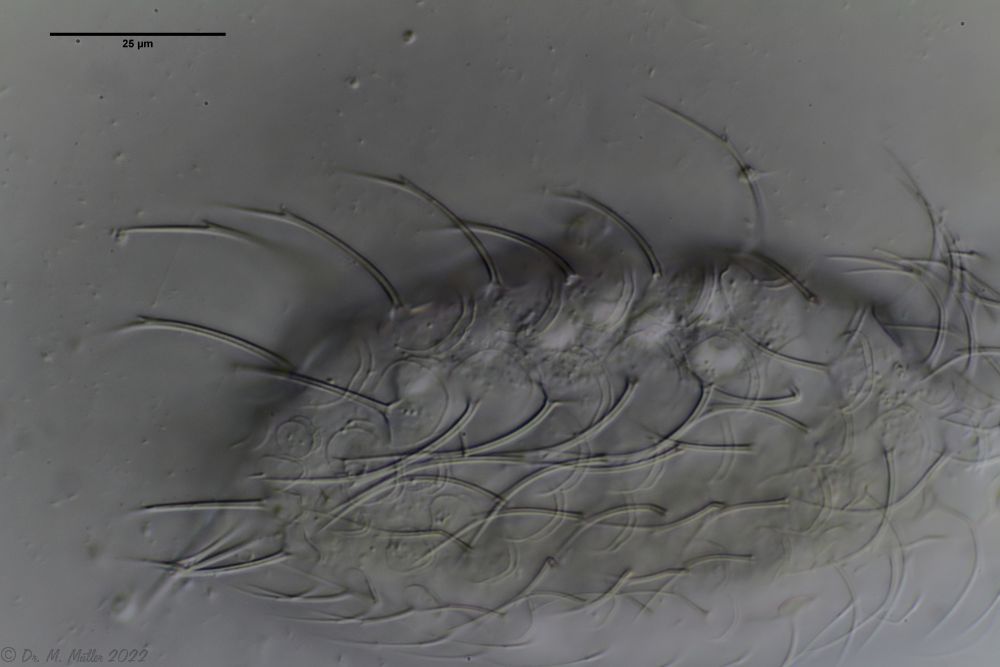
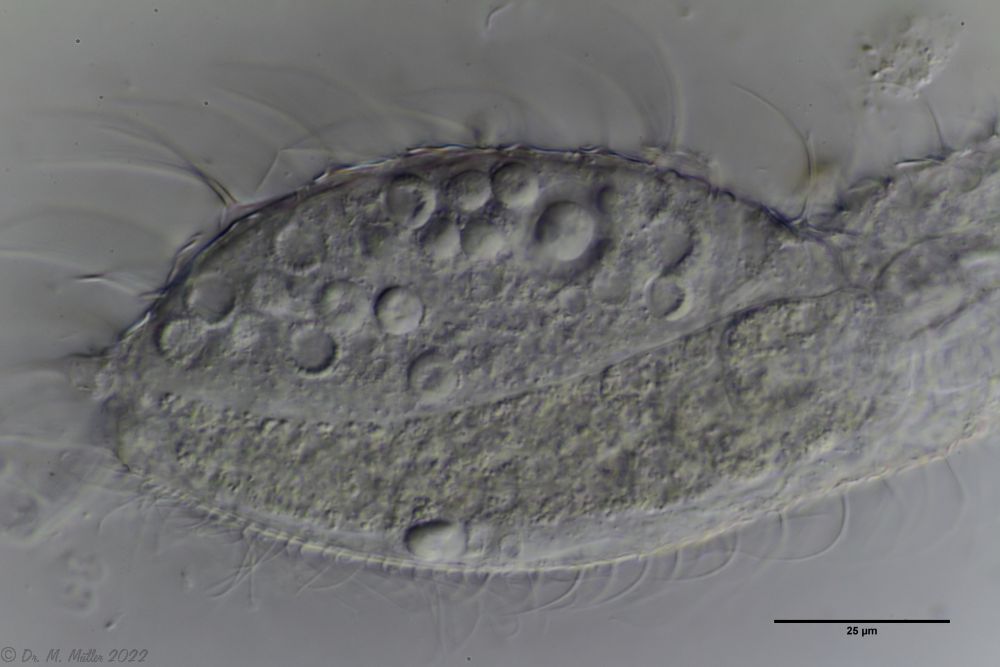
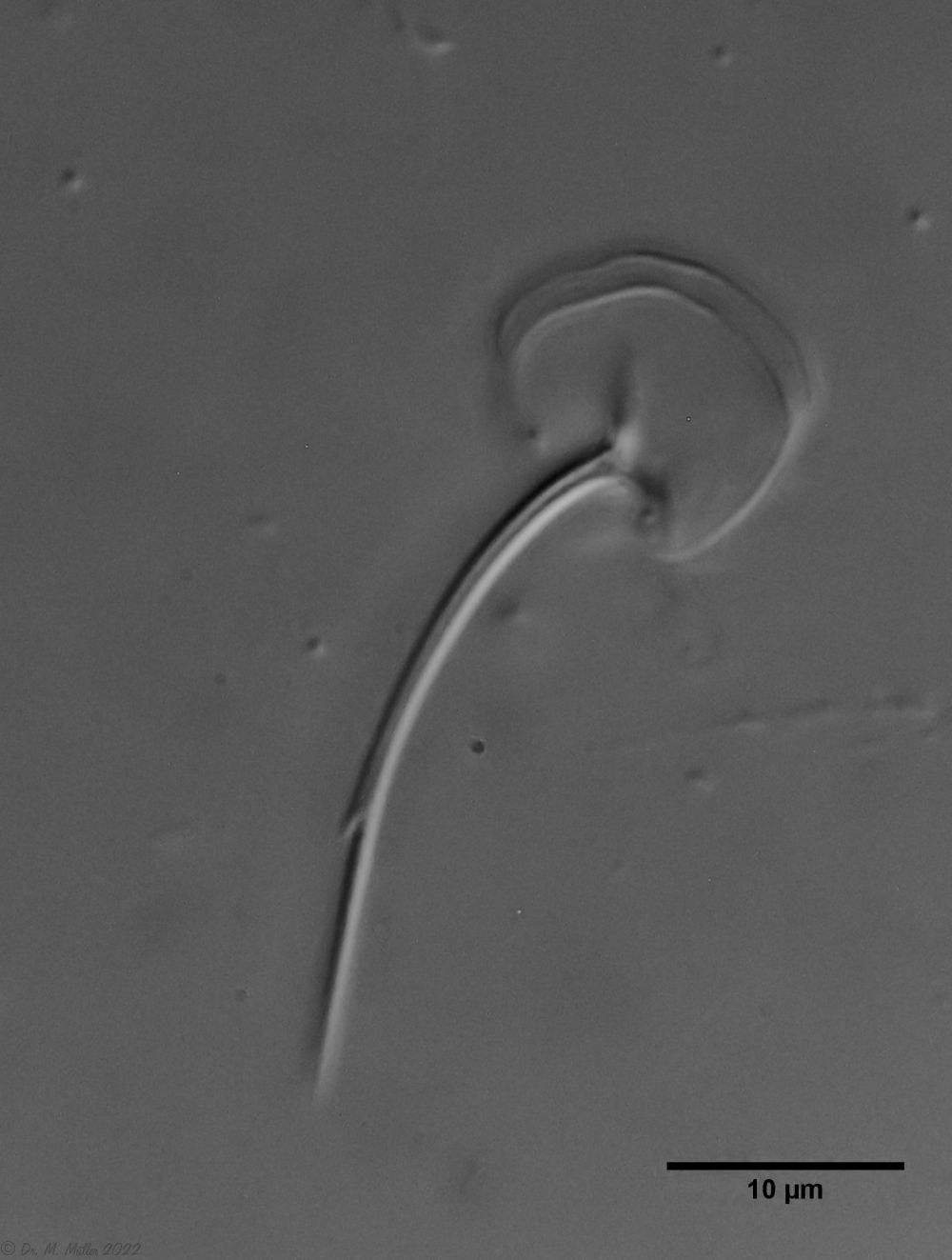 Ch. cordiformis: herzförmige Schuppen
Ch. cordiformis: herzförmige Schuppen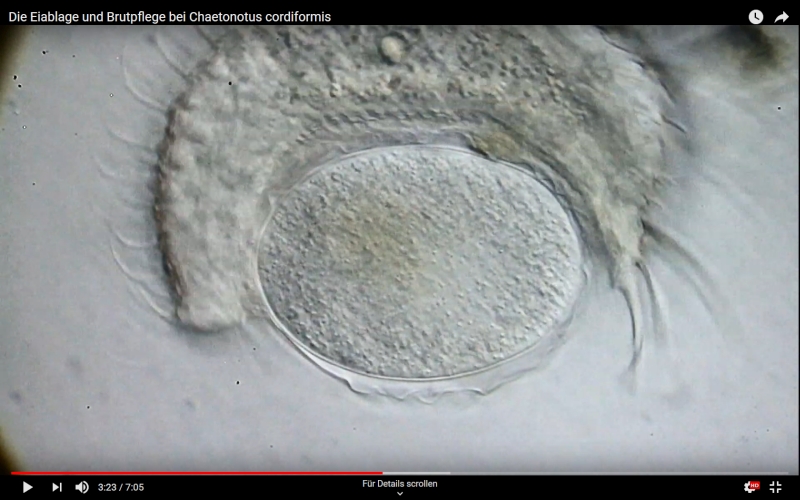
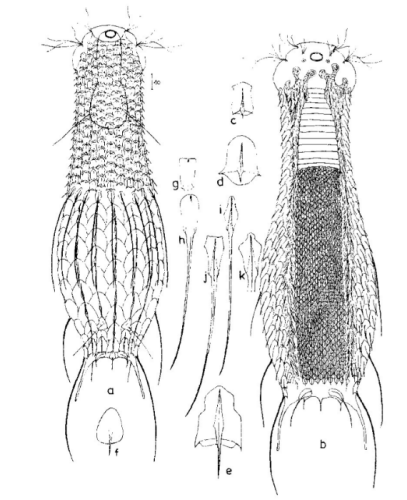
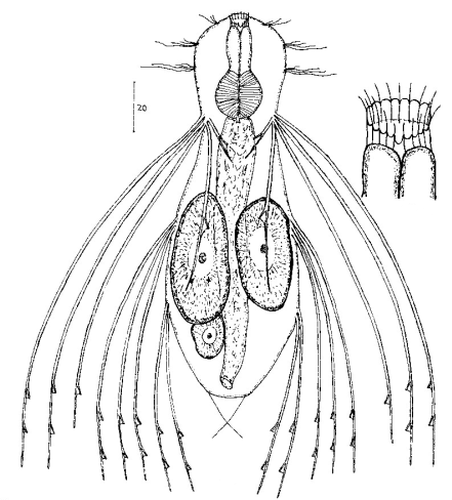
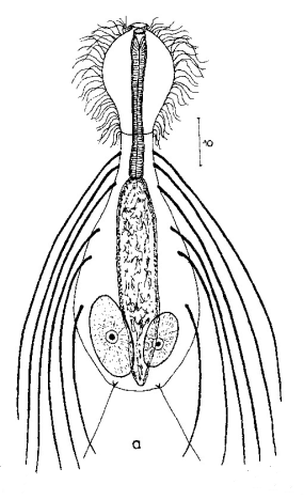
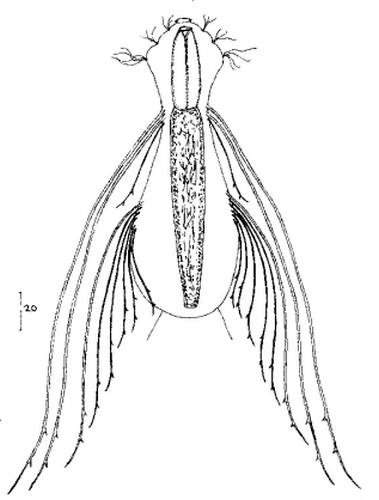
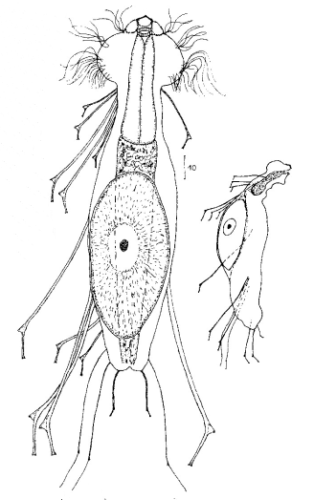
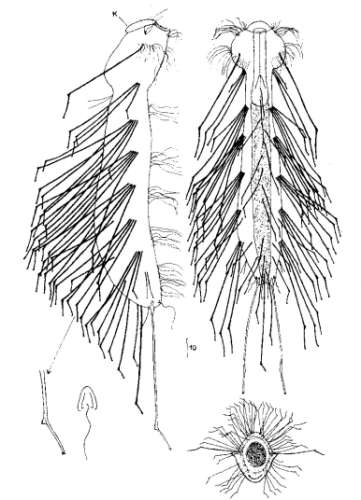
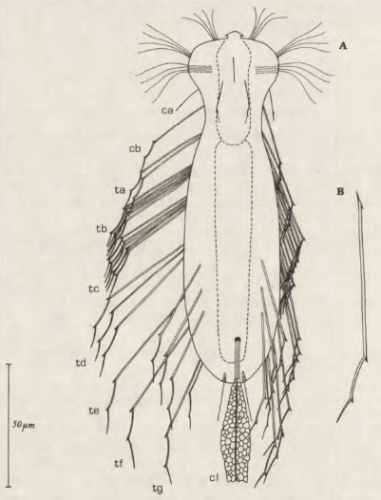
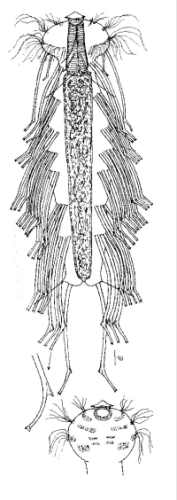
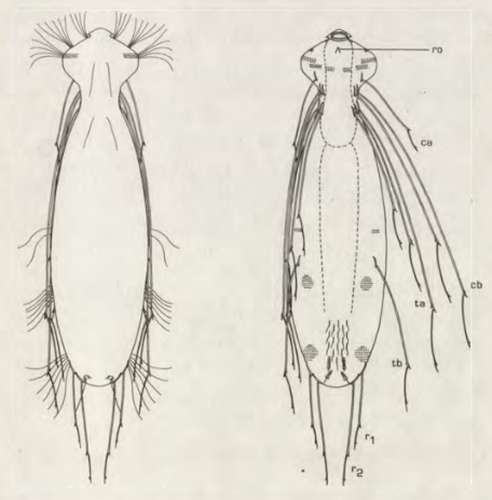
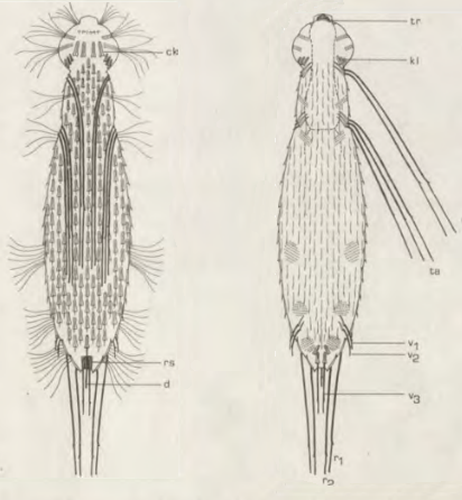
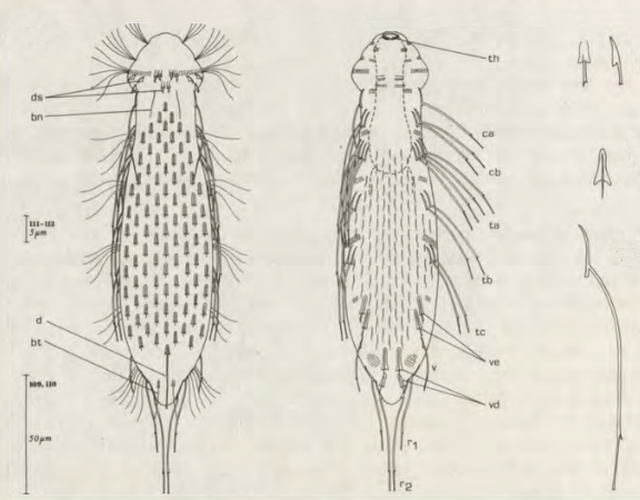
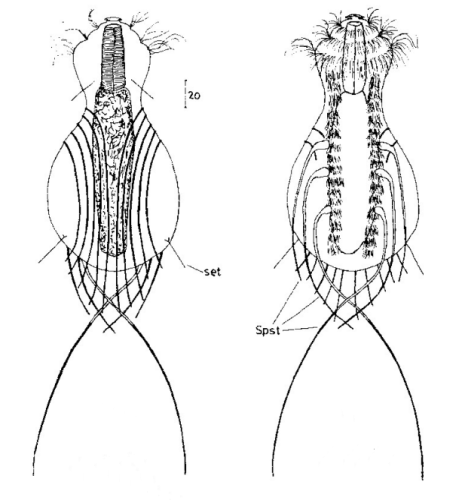
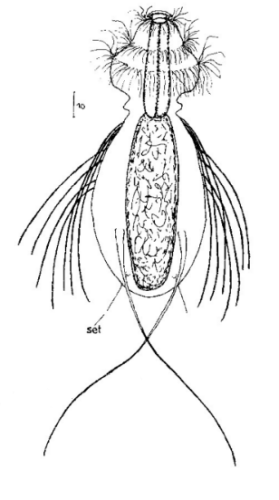
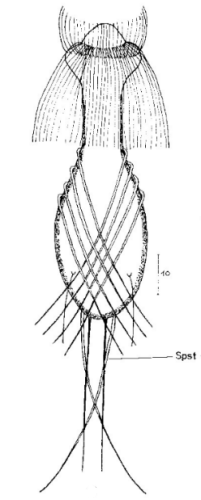
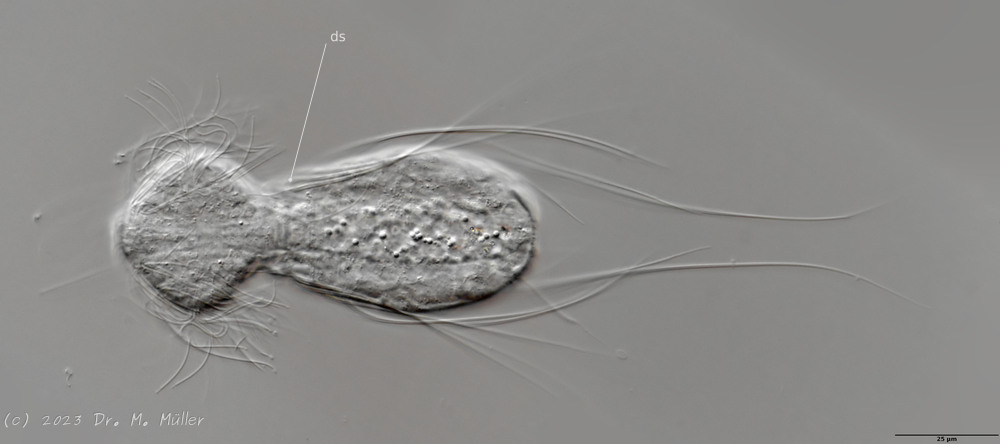 Dorsal; ds: dorsale Stacheln (5 Paar)
Dorsal; ds: dorsale Stacheln (5 Paar)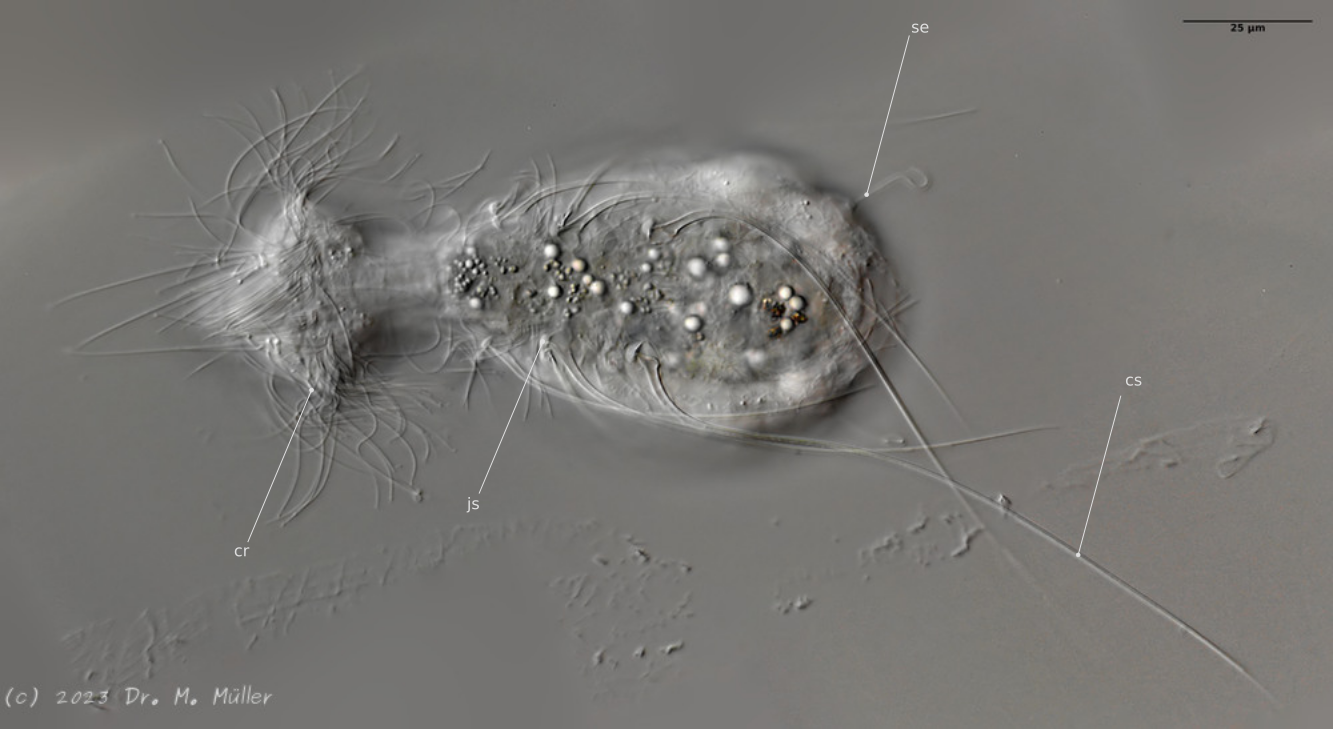
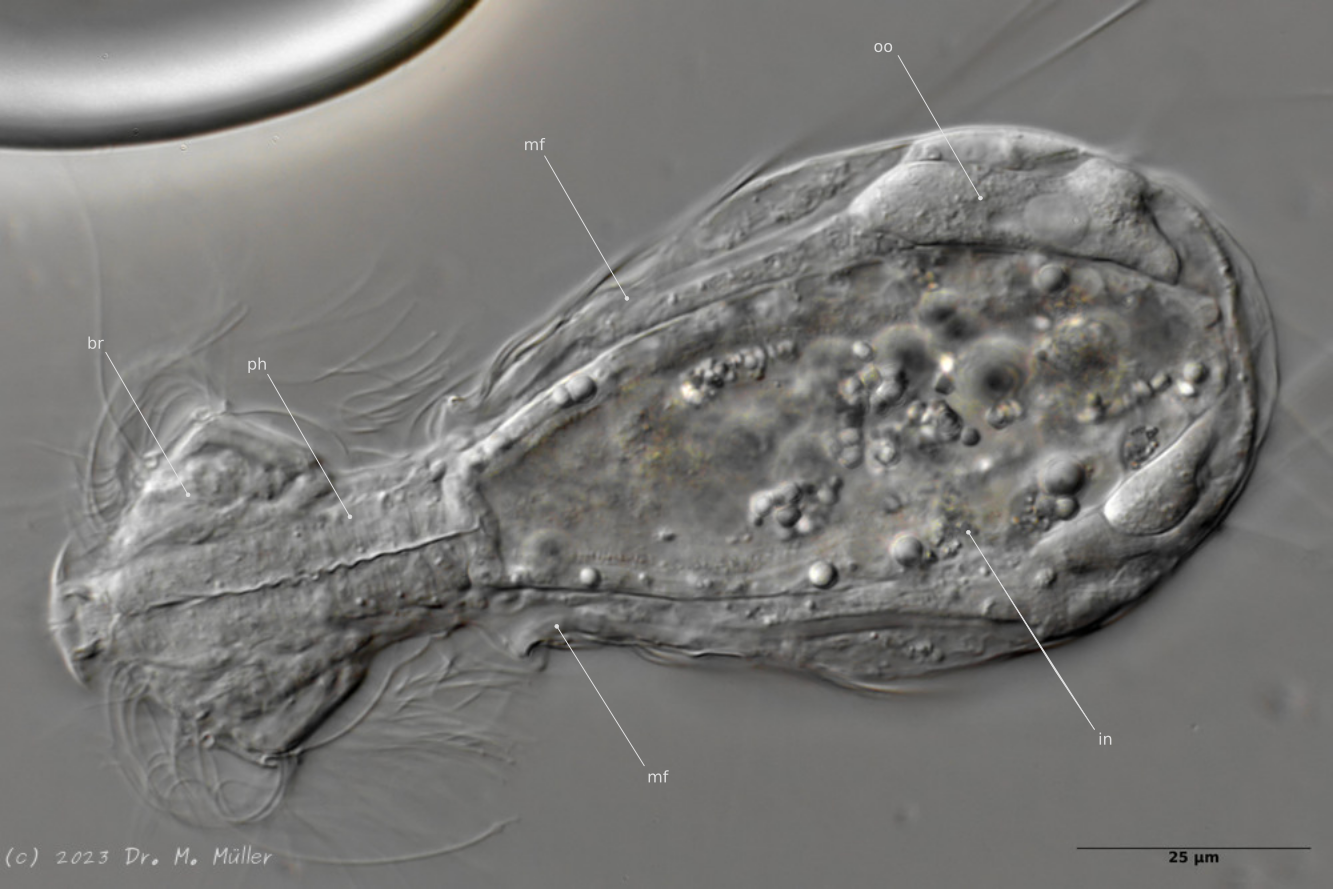
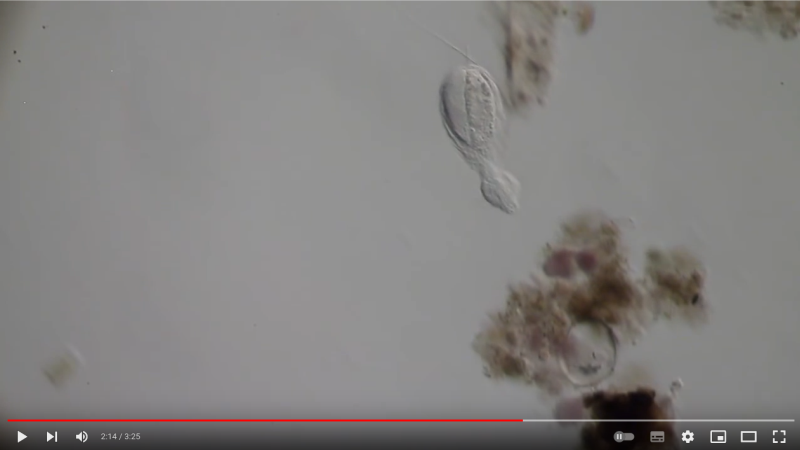
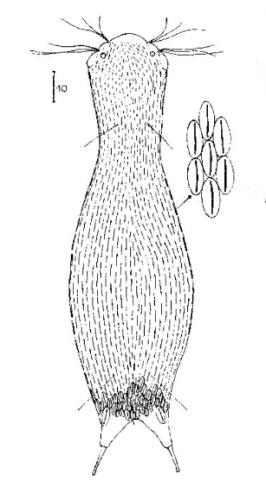
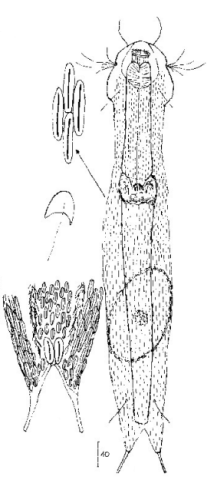
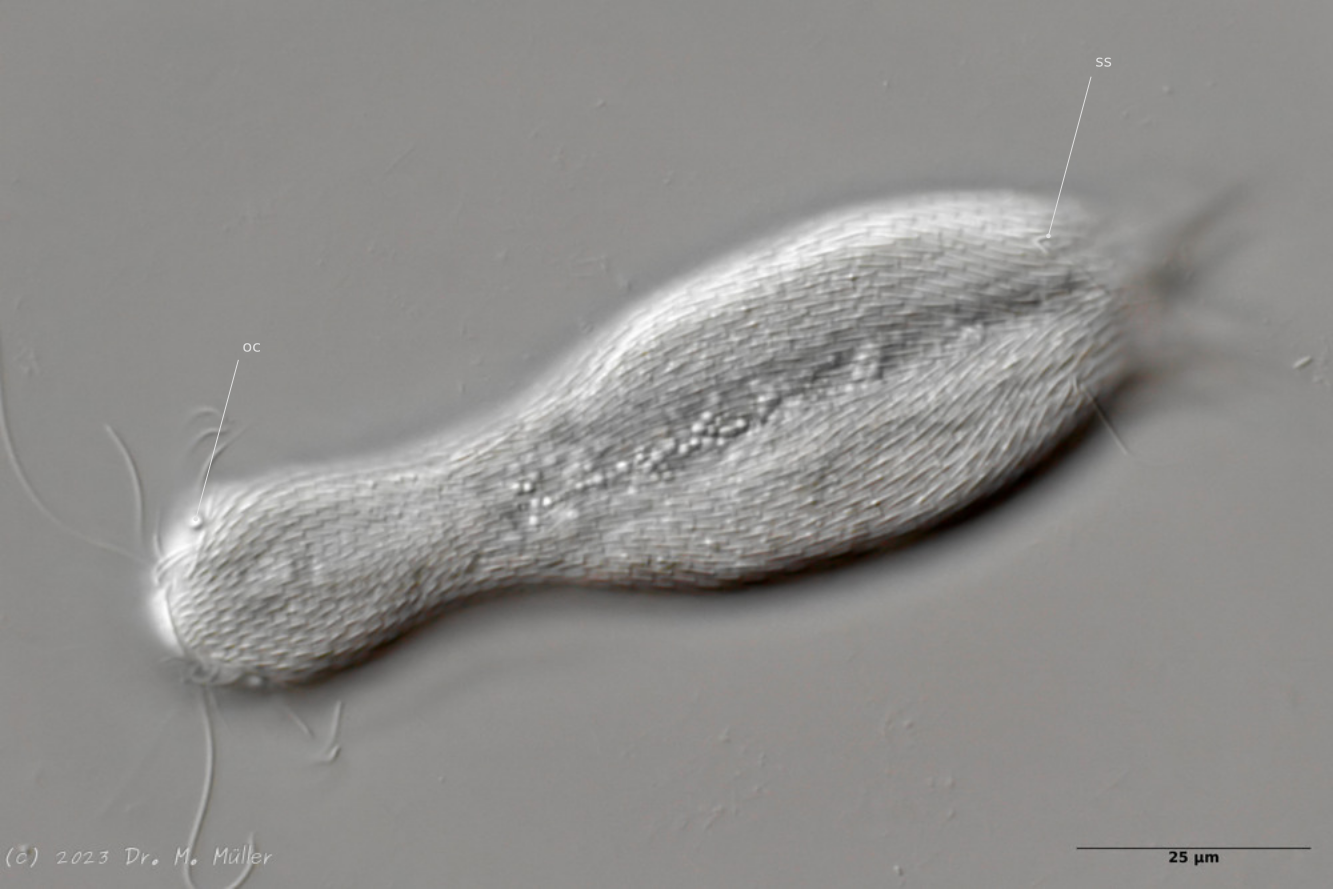
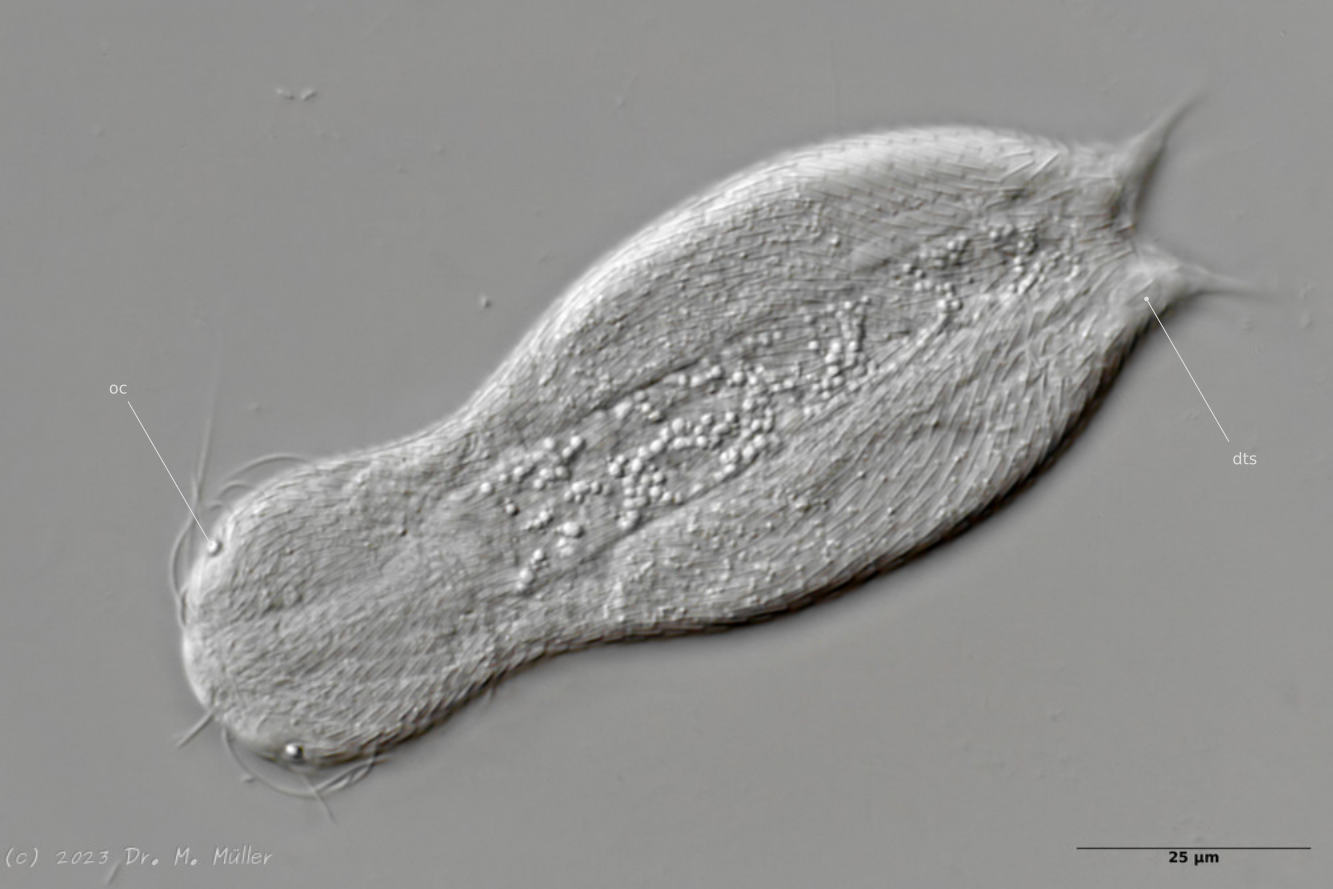
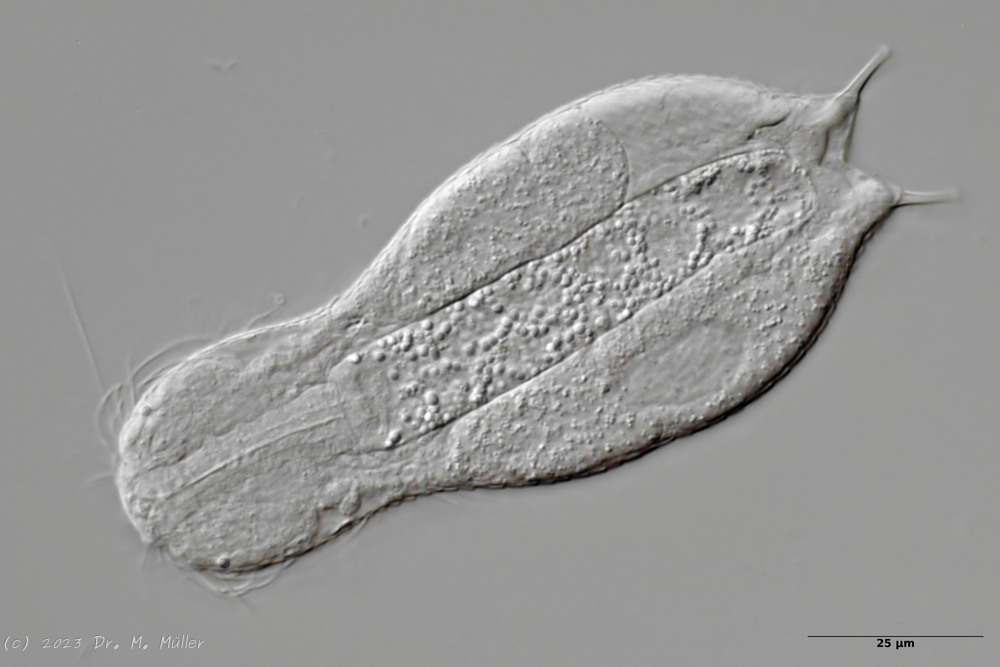
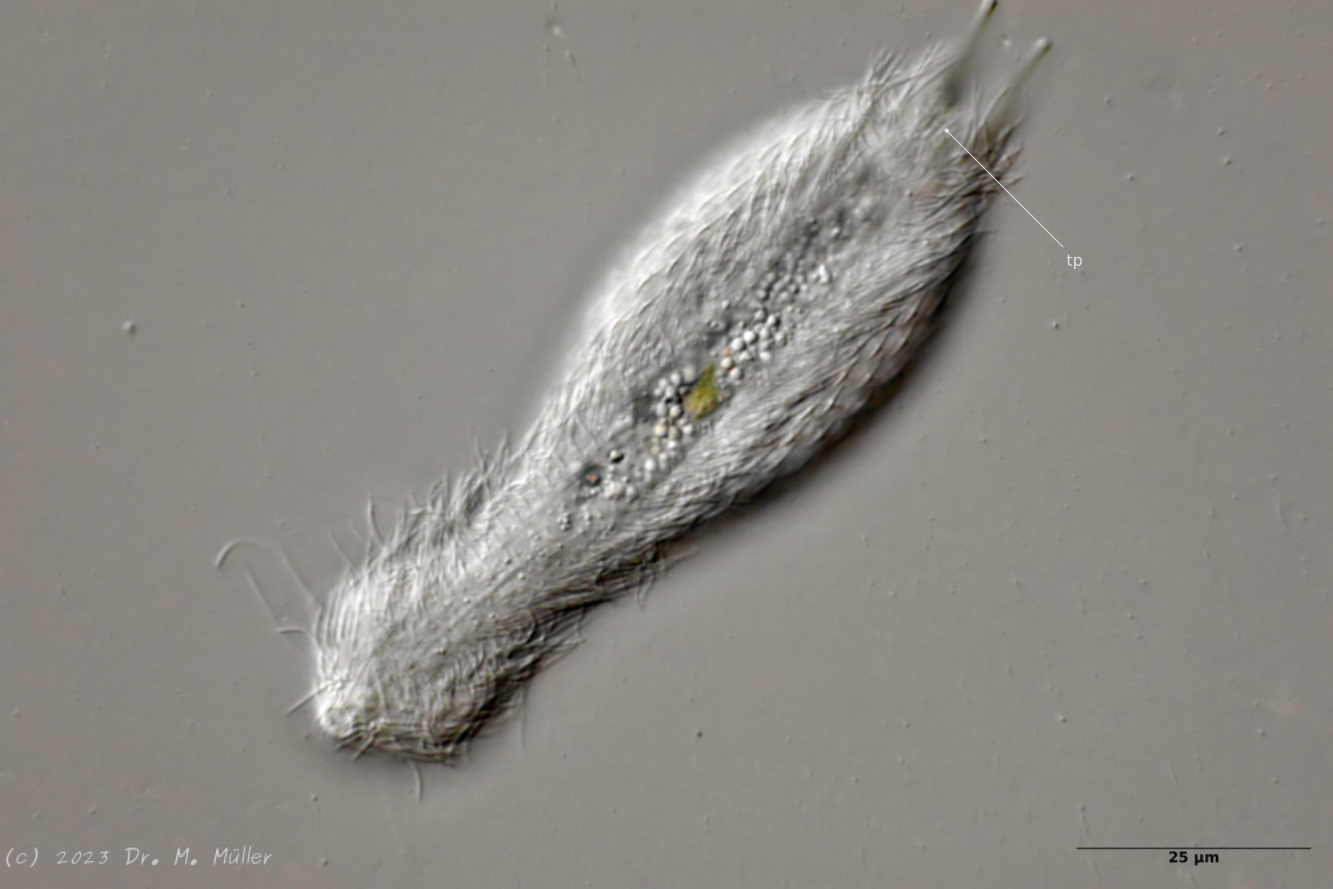
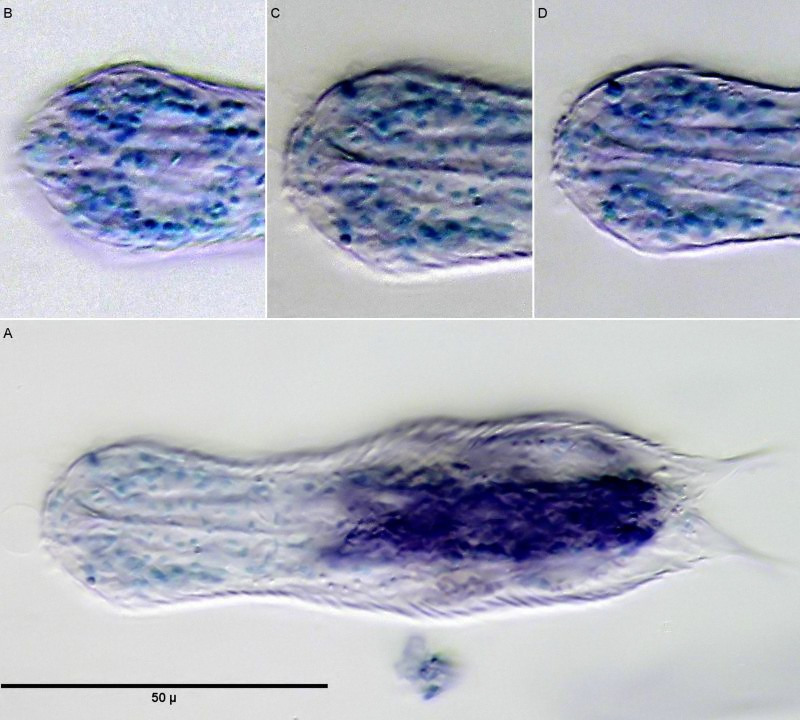
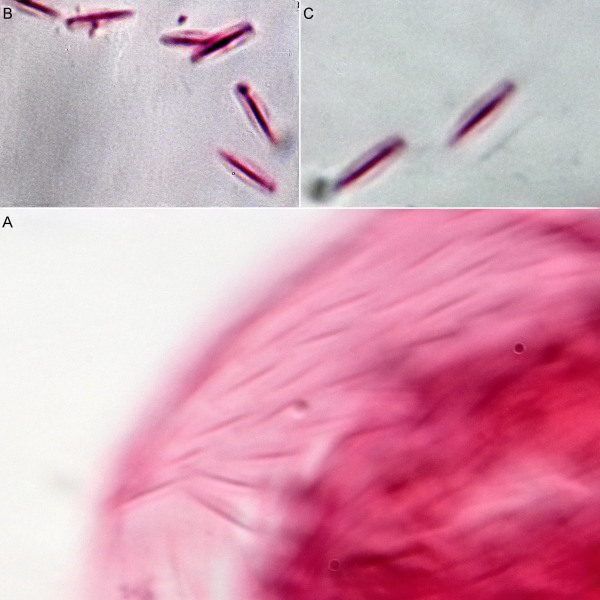
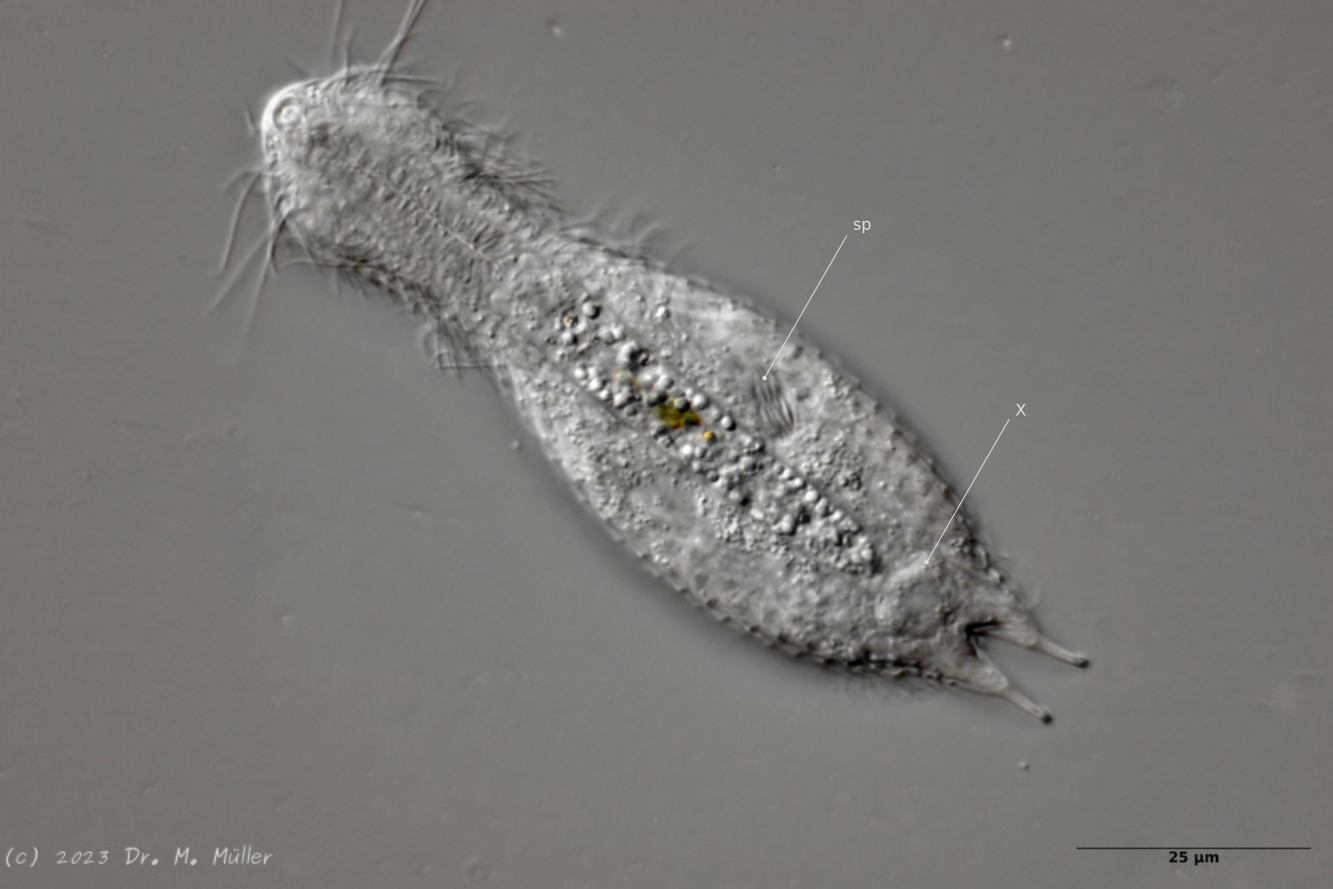
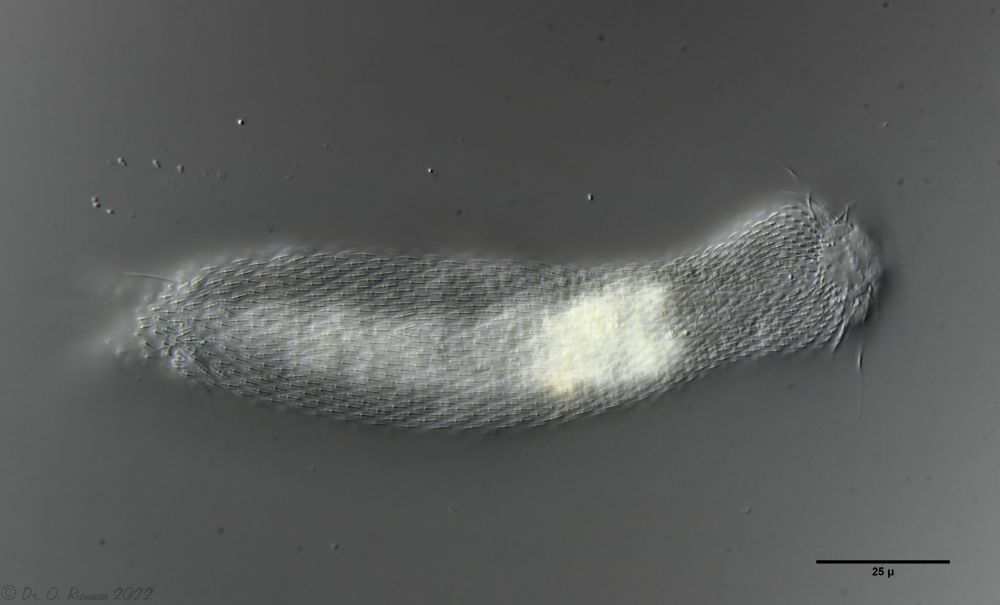
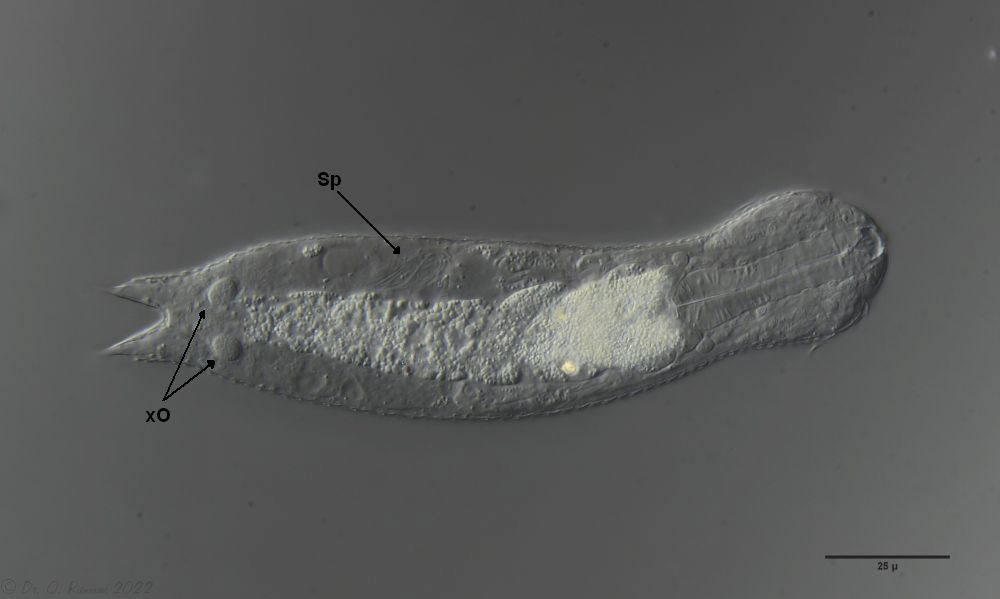
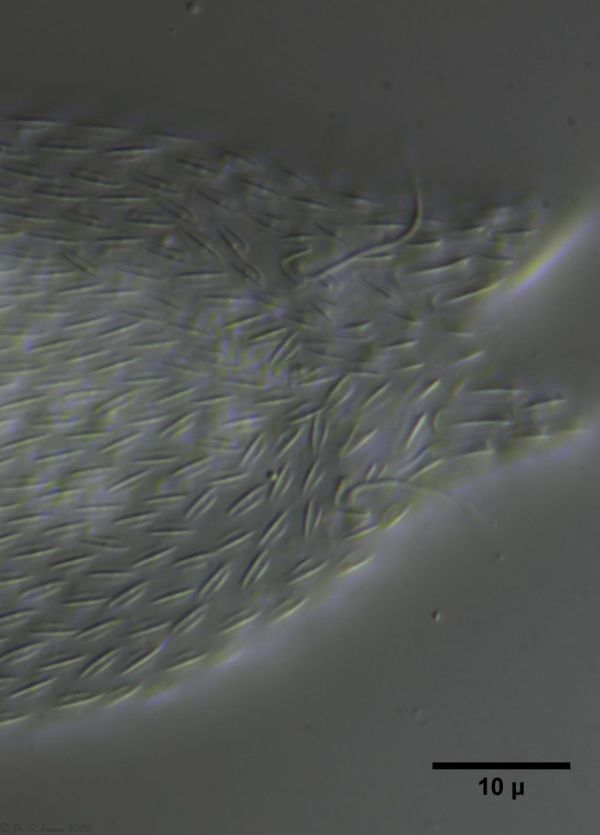
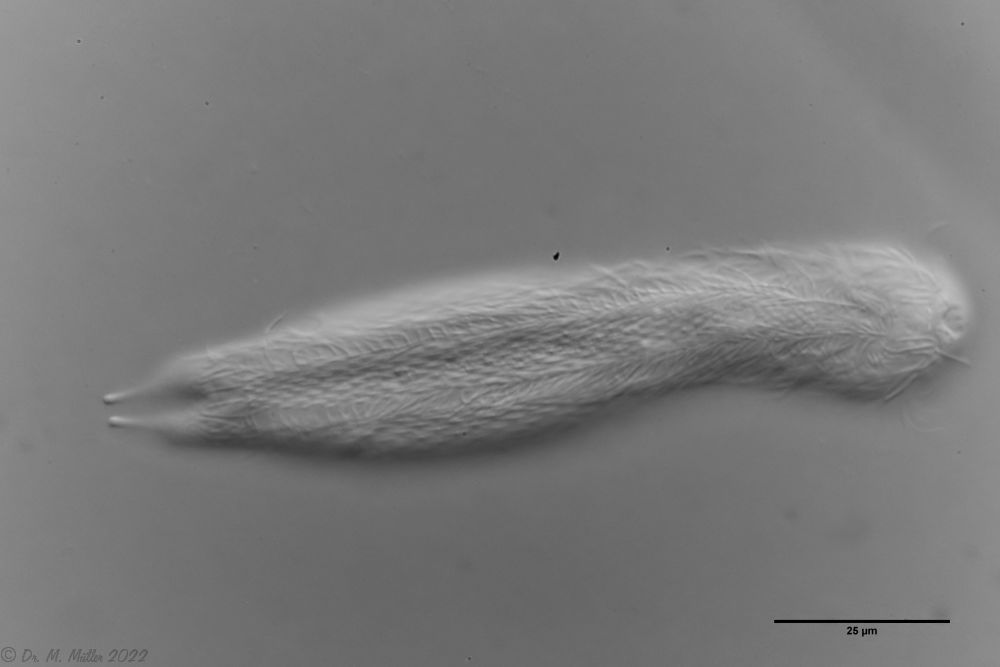
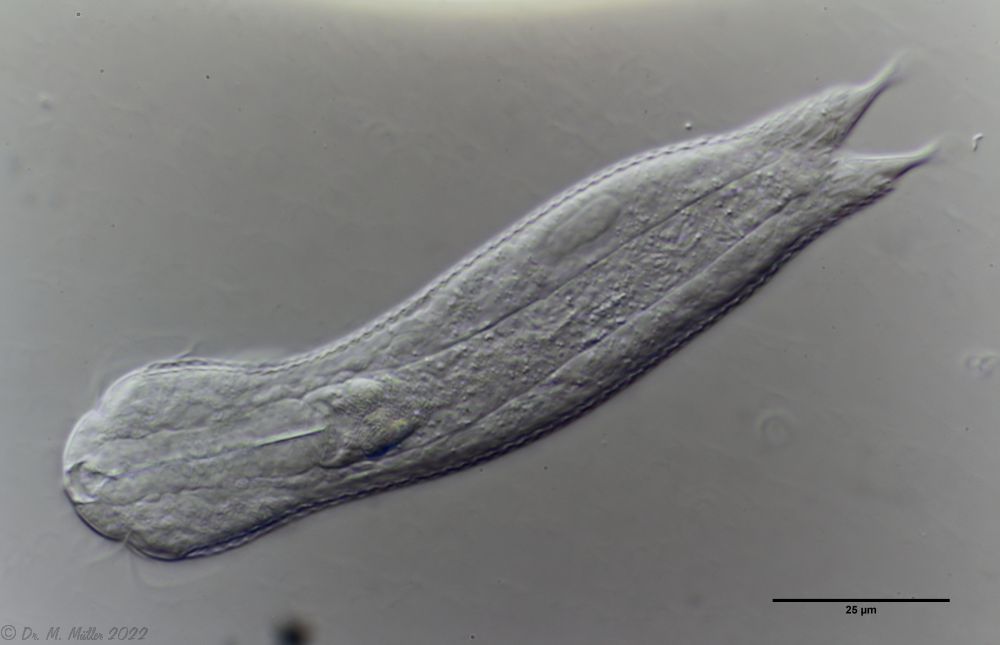
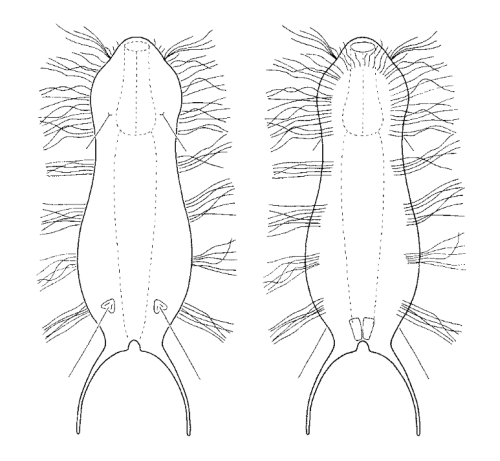
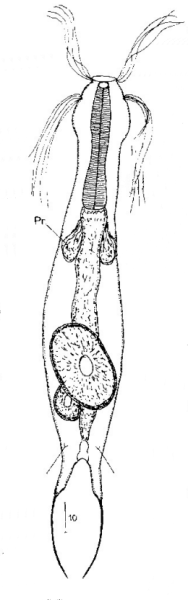
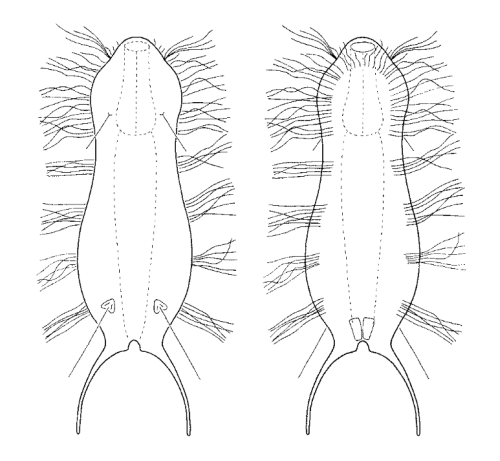
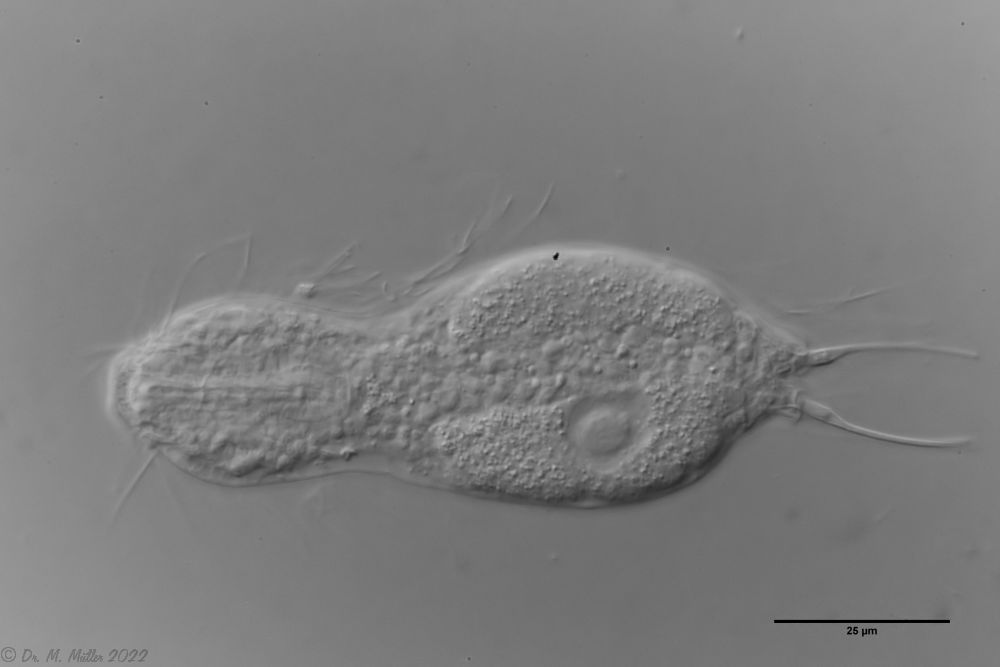
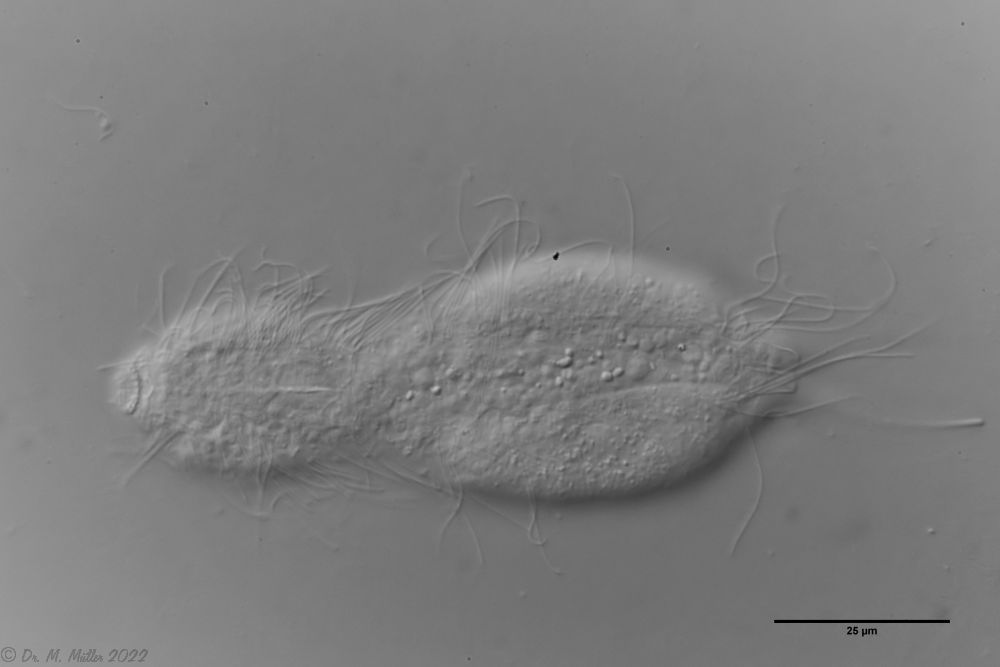
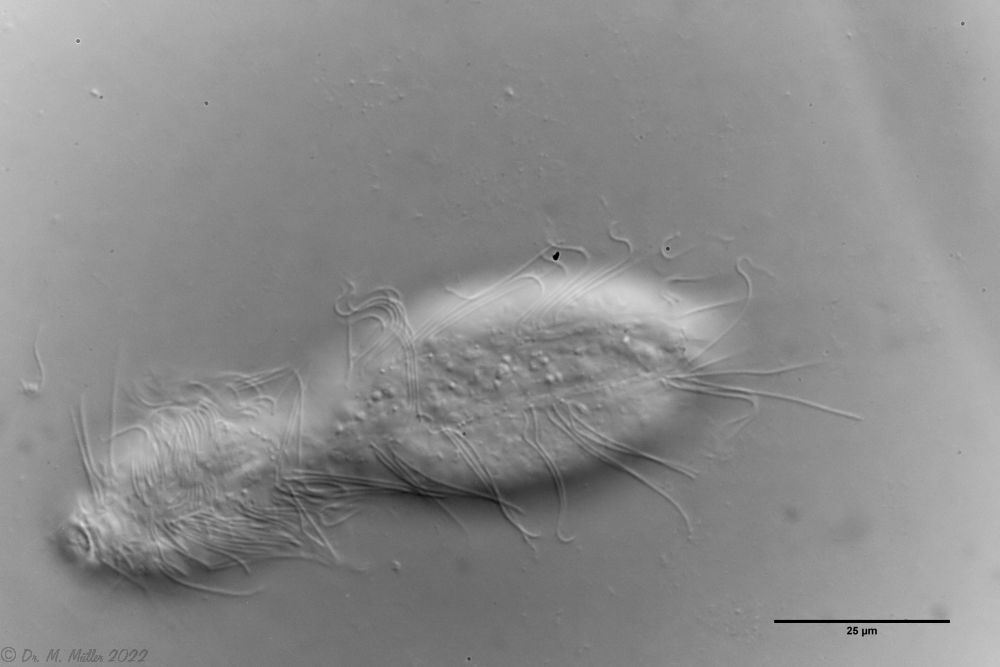
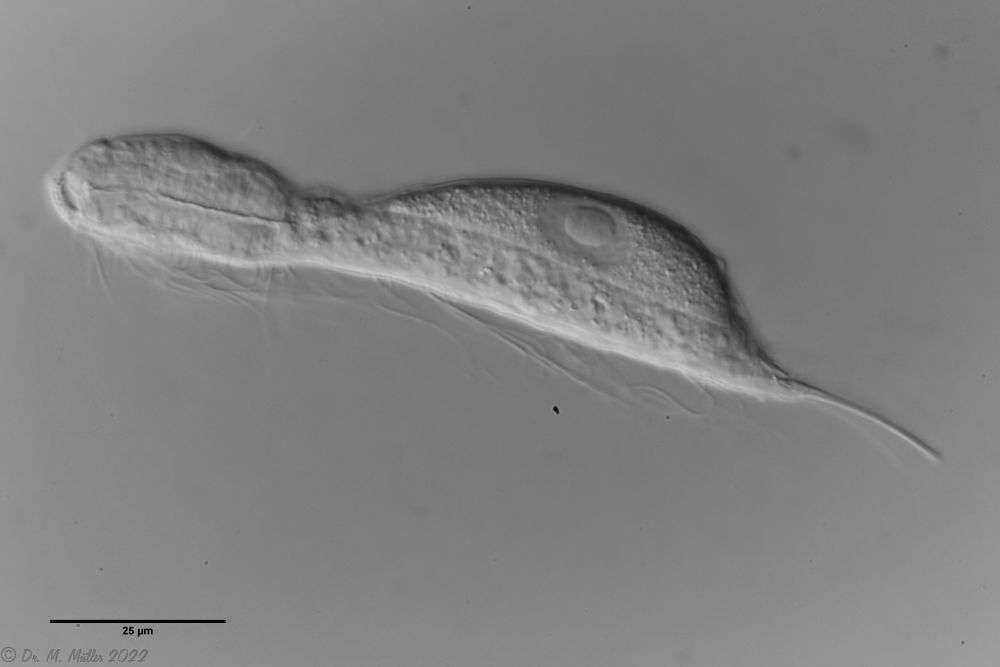
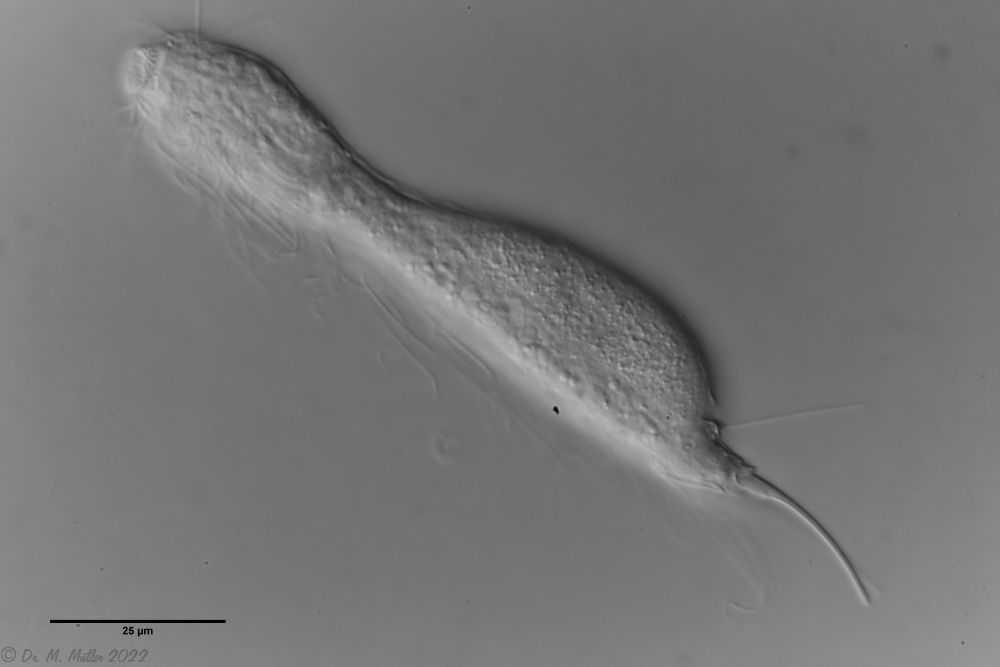
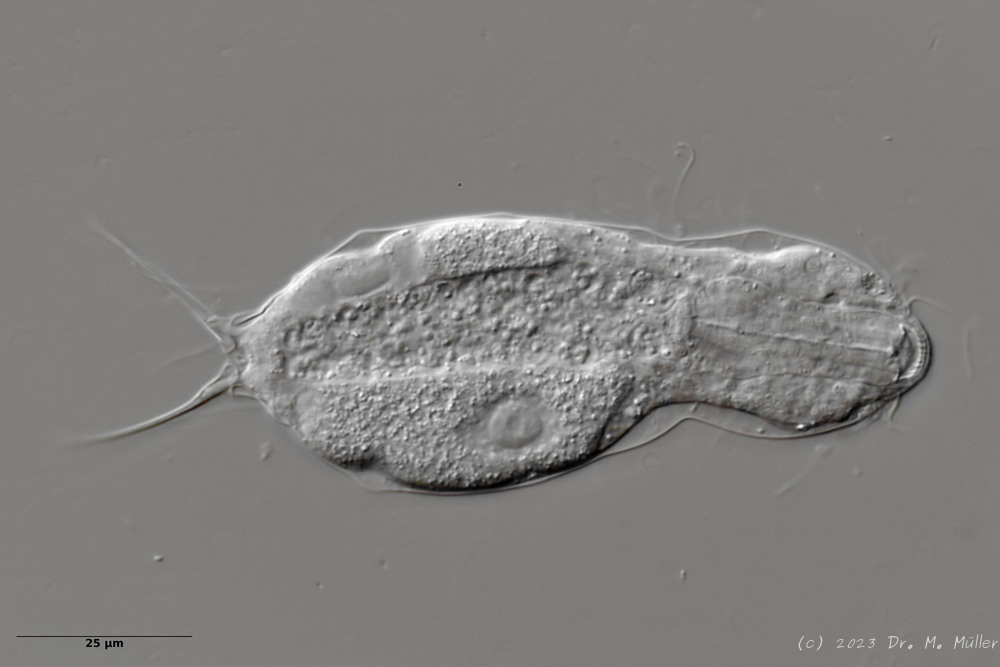
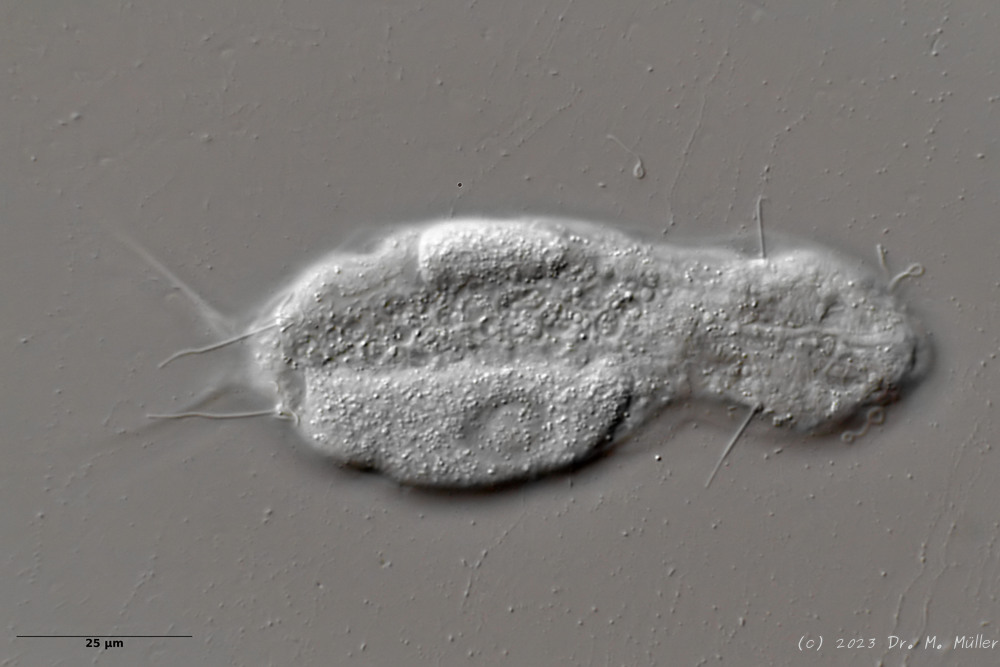
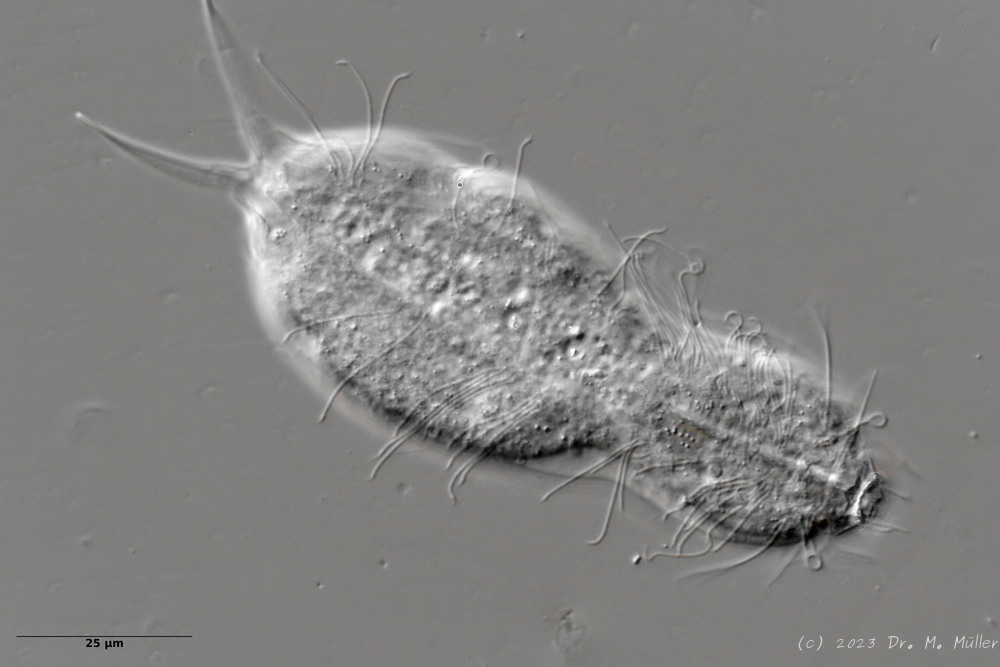

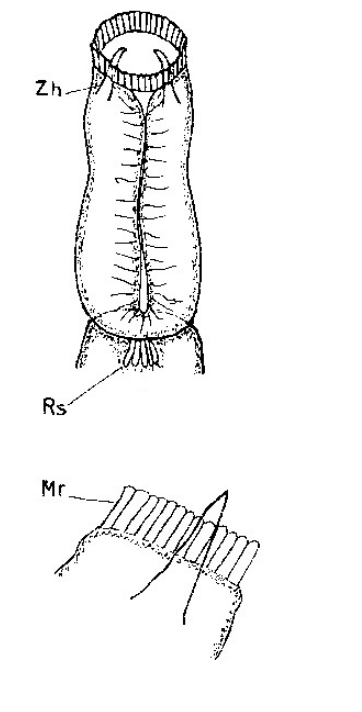
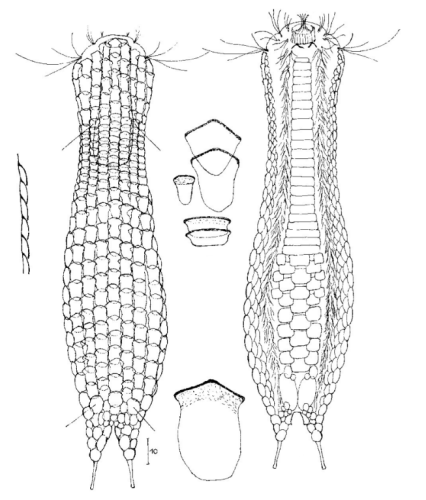
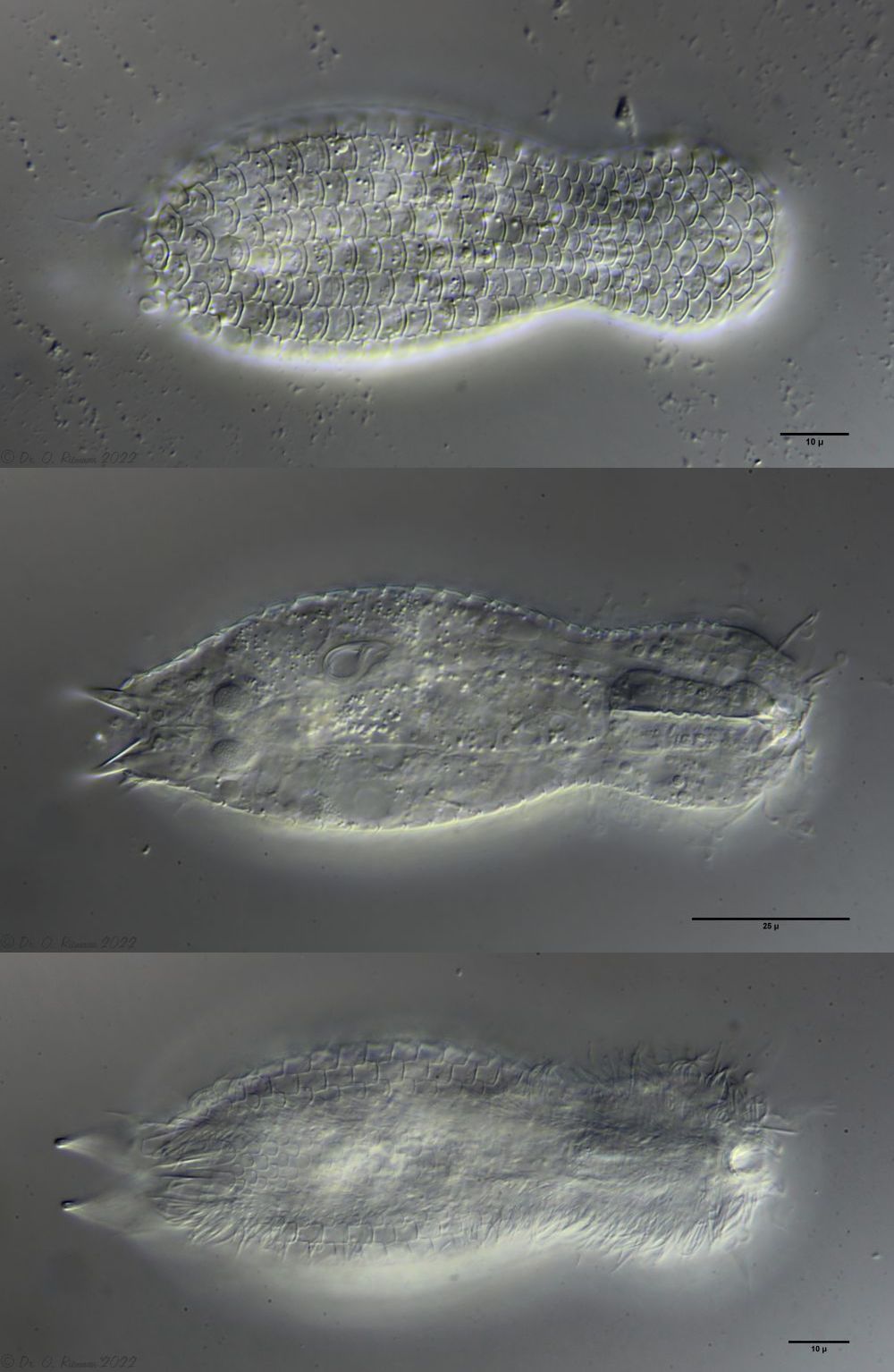 L. minor chaetifer: oben dorsal mit den für Lepidodermella typischen glatten Schuppen;
L. minor chaetifer: oben dorsal mit den für Lepidodermella typischen glatten Schuppen;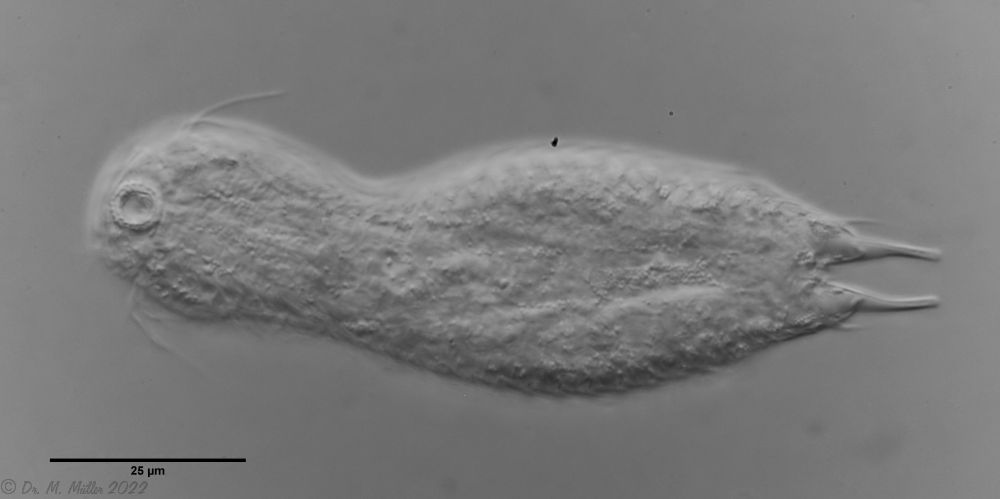 L. minor chaetifer: Das Stachelpaar an der Zehenbasis ist charakteristisch für die Unterart chaetifer
L. minor chaetifer: Das Stachelpaar an der Zehenbasis ist charakteristisch für die Unterart chaetifer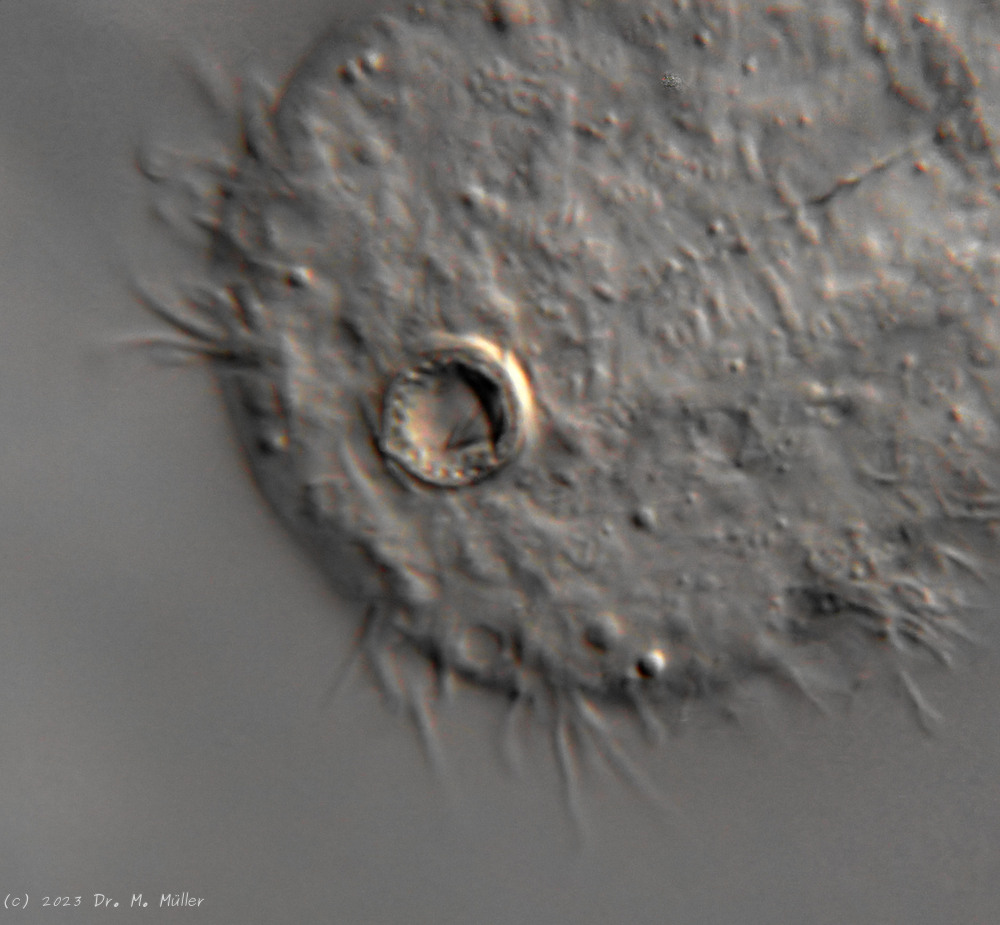
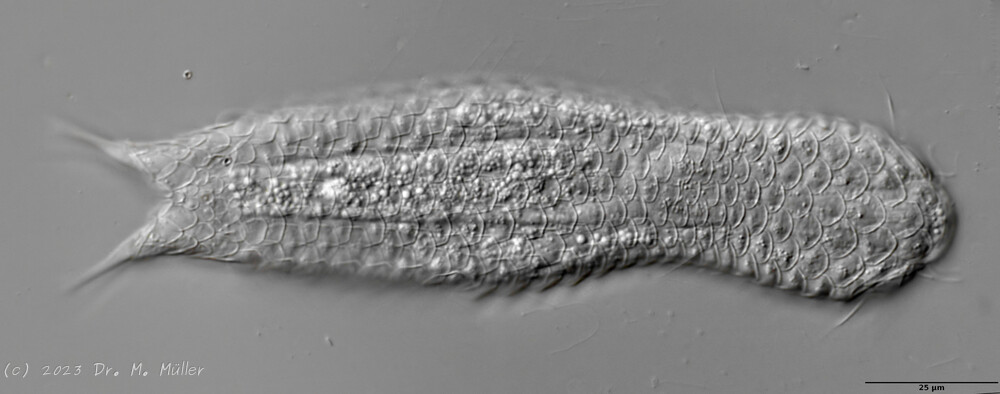 Dorsale Schuppen
Dorsale Schuppen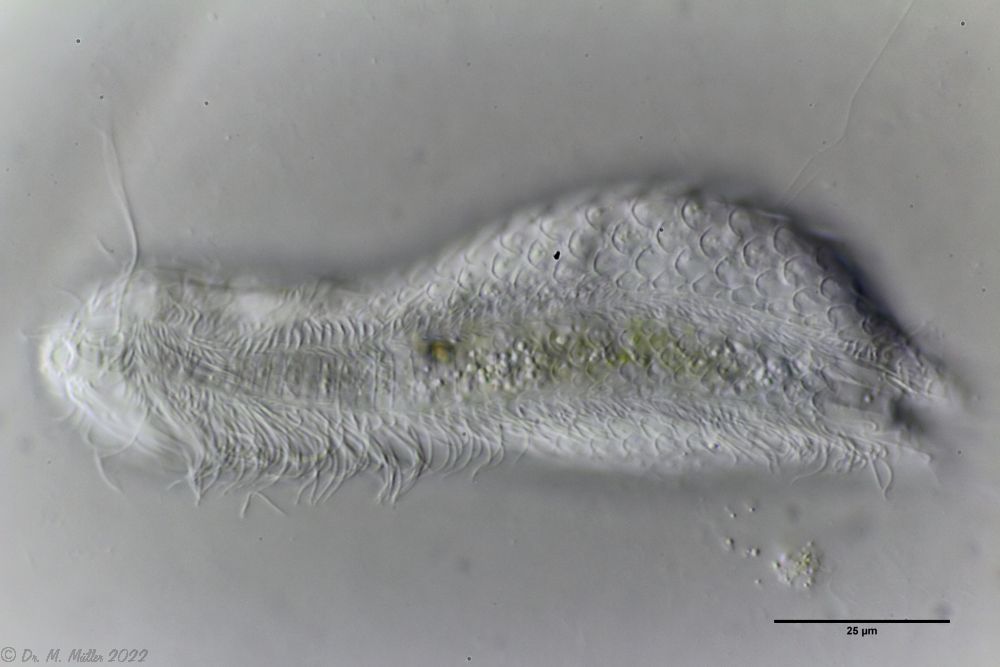 Ventrale Schuppen; große Terminalplatten; rechteckige Spangen unter Hals
Ventrale Schuppen; große Terminalplatten; rechteckige Spangen unter Hals